Тема 4
Эмбриогенез анамний
1. Общая характеристика анамний и амниот.
2. Эмбриогенез анамний.
3. Эмбриогенез ланцетника.
4. Эмбриогенез земноводных, миног.
5. Эмбриогенез хрящевых и костистых рыб.
1. Антипчук, Ю.П. Гистология с основами эмбриологии / Ю.П. Антипчук. – М.: Просвещение, 1983. – 240 с.
2. Алмазов, И.В., Сутулов Л.С. Атлас по гистологии и эмбриологии / И.В. Алмазов, Л.С. Сутулов. – М.: Медицина, 1978. – 148 с.
3. Гистология / под ред. Ю.И. Афанасьева. – М: Медицина, 1989. – 361 с.
4. Рябов, К.П. Гистология с основами эмбриологии / К.П. Рябов. – Мн.: Высш. шк., 1991. – 289 с.
5. Биологический энциклопкдический словарь / под ред. М.С. Гилярова. – М.: Сов. Энцикл., 1989. – 864 с.
6. Практикум по гистологии, цитологии и эмбриологии / под ред. Н.А. Юриной, А.И. Радостиной. – М.: Высш. шк., 1989. – 154 с.
Хэм А., Кормик Д. Гистология / А. Хэм, Д. Кормик. – М.: Мир, 1983. – 192
1. Особенности эмбрионального развития млекопитающих.
2. Эмбриогенез яйцекладущих млекопитающих.
3. Эмбриогенез сумчатых млекопитающих.
4. Эмбриогенез плацентарных млекопитающих.
5. Эмбриогенез человека.
Общая характеристика анамний и амниот
Общая характеристика анамний
Исходя из особенностей эмбрионального развития, все хордовые подразделяются на две группы: анамнии и амниоты. Анамнии – это животные, у которых в процессе эмбрионального развития не образуются такие зародышевые оболочки, как амнион, или водная оболочка, и аллантоис. К анамниям относят хордовых, ведущих первичноводный образ жизни, а также низших хордовых, тесно связанных с водной средой в период размножения и эмбрионального развития зародышей – бесчелюстных, рыб и земноводных. В связи с эмбриональным развитием этих хордовых в водной среде, у них отсутствует водная оболочка и аллантоис, так как функции дыхания, выделения и питания развивающегося зародыша обеспечивает окружающая его водная среда.
Хордовых, относящихся к анамниям по характеру эмбрионального развития можно подразделить на три группы:
1) ланцетник, яйцеклетки которого содержит мало желтка;
2) некоторые круглоротые, рыбы (хрящевые ганоиды) и земноводные, яйцеклетки которых содержат среднее количество желтка;
3) селяхии и костистые рыбы, яйцеклетки содержат много желтка.
Эмбриогенез ланцетника
После оплодотворения в яйцеклетке ланцетника начинается перераспределение желтка, который концентрируется в основном на одной стороне яйцеклетки, соответствующей вегетативному полюсу. Анимальный полюс яйцеклетки определяется по расположенному над ним второму полярному тельцу. Дробление яйцеклетки полное, равномерное (рисунок 1).
/ – анимальный полюс; 2 – вегетативный полюс; 3 –скопление желтка; 4 –целобластула; 5 – клетки бластодермы.
Рисунок –1. Последовательность (I –VI) дробления яйцеклетки ланцетника
Первые два дробления идут меридионально, третье – экваториально. Дальнейшее дробление идет попеременно то в одном, то в другом направлении, и количество клеток увеличивается в геометрической прогрессии. После образования однослойного зародыша – бластулы становится заметным, что клетки анимального полюса более мелкие, чем клетки вегетативного полюса. В шарообразной целобластуле ланцетника различают уплощенную часть вегетативного полюса, называемую дном бластулы, а противоположную часть, соответствующую анимальному полюсу, называют крышей бластулы. Клетки, образующие крышу бластулы, будут дифференцироваться в клетки наружного зародышевого листка, или эктодерму, а клетки дна бластулы – в энтодерму.
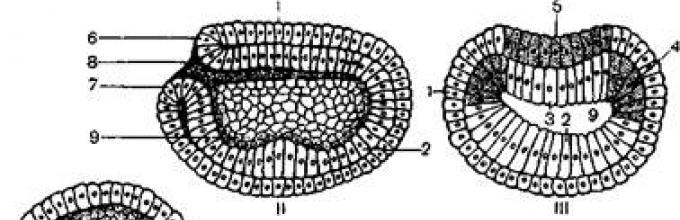
Гаструляция осуществляется путем инвагинации бластодермы вегетативного полюса внутрь бластоцеля. Впячивание продолжается до тех пор, пока клетки вегетативного полюса соприкоснутся с клетками анимального полюса, в связи с чем полость бластоцеля суживается и исчезает (рисунок 2).

I– целобластула; II – IV – гаструляция; V–нейрула;
1 – эктодерма; 2–энтодерма; 3 –хорда; 4–мезодерма; 5 – нервная пластинка; 6 –верхняя и 7 – нижняя губа бластопора; 8 – бластопор; 9–полость первичной кишки; 10– полость вторичной кишки; 11 – целом.
Рисунок 2 –Эмбриогенез ланцетника
С завершением первого этапа гаструляции возникает двуслойный зародыш, или гаструла, состоящий из клеток наружного зародышевого листка – эктодермы и внутреннего зародышевого листка – энтодермы. В результате впячивания образуется полость первичной кишки, выстланная клетками энтодермы, которая сообщается с внешней средой бластопором. Клеточный состав энтодермы неоднородный, поскольку в нее входит также клеточный материал будущей хорды и мезодермы. С образованием полости первичной кишки зародыш начинает быстро расти и удлиняется, но наиболее интенсивные формообразовательные процессы осуществляются в области верхней, или дорсальной, губы бластопора. Непосредственно за верхней губой бластопора, на спинной поверхности зародыша, эктодерма утолщается и состоит из высоких призматических клеток, называемых медуллярной или нервной пластинкой. Эктодерма, окружающая нервную пластинку, представлена мелкими клетками, которые образуют кожный покров. Под нервной пластинкой такие же изменения претерпевают клетки энтодермы, которые представляют материал будущей хорды. В дальнейшем нервная пластинка начинает прогибаться, образуя нервный желобок, а клетки кожной эктодермы интенсивно наползают на него. Впоследствии нервный желобок углубляется, края его смыкаются, и он превращается в нервную трубку, полость которой именуется, нервным каналом. Клетки кожной эктодермы смыкаются, и нервная трубка оказывается под ними. Одновременно клетки энтодермы, примыкающие к нервной пластинке, прогибаются в сторону последней, скручиваются и обособляются в плотный тяж – хорду, которая имеет вид сплошного цилиндра. По сторонам от хордального зачатка энтодерма инвагинирует в сторону эктодермы, образуя мезодермальные выпячивания, или мезодермальные мешки, которые впоследствии отшнуровываются от энтодермы и начинают разрастаться между эктодермой и энтодермой. Полость мезодермальных мешков, возникающая из гастроцеля, превращается во вторичную полость тела, или целом. Таким образом, в процессе гаструляции возникает трехслойный зародыш.
После обособления хорды и отшнуровывания мезодермальных мешков края энтодермы постепенно сближаются в спинной части зародыша и, смыкаясь, образуют замкнутую кишечную трубку. Вслед за гаструляцией у зародыша возникает комплекс осевых органов, характерный для представителей типа хордовых. Он состоит из хорды, по бокам которой располагаются скопления сегментированной мезодермы – сомиты.
Закладка осевых органов происходит на стадии нейрулы. Нервная трубка ланцетника в передней и задней частях зародыша некоторое время остается открытой. В дальнейшем на задней части тела зародыша эктодерма нарастает на бластопор и закрывает его так, что полость нервной трубки сообщается с кишечной полостью нервно-кишечным каналом, который быстро зарастает. Ротовое отверстие у зародыша ланцетника образуется вторично на переднем конце тела вследствие истончения и прорыва эктодермы.
Третий зародышевый листок, или мезодерма, зародыша ланцетника сегментирована на всем протяжении. Мезодермальные сегменты в дальнейшем разделяются на спинную часть – сомиты и брюшную часть – спланхнотомы. Сомиты остаются сегментированными, а спланхнотомы на каждой стороне тела утрачивают первичную сегментацию, сливаются и образуют, расщепляясь на два листка, правую и левую целомические полости. Последние объединяются под кишечной трубкой в общую вторичную полость тела. Когда у ланцетника начинает формироваться хвост, то нервно-кишечный канал исчезает, а на заднем конце зародыша на месте бластопора вследствие истончения и прорыва стенки тела возникает анальное отверстие. Пройдя описанные стадии развития, ланцетник становится свободноплавающей личинкой. В период личиночного развития завершается органогенез и гистогенез и личинка превращается во взрослое животное.
Индивидуальное развитие ланцетника представляет собой простейшую исходную схему эмбриогенеза, путем постепенного усложнения которой в ходе эволюции возникли более сложные системы развития хордовых животных, включая человека.
СТРОЕНИЕ ЯЙЦЕКЛЕТКИ. ОПЛОДОТВОРЕНИЕ
Яйца ланцетника бедны желтком и микроскопически малы (100-120мкм), относятся к изолецитальному типу. Желточные зерна невелики и распределены в цитоплазме почти равномерно. Тем не менее, в яйцеклетке выделяют анимальный и вегетативный полюс. В области анимального полюса при созревании яйцеклетки происходит отделение редукционных телец. Ядро в оплодотворенной яйцеклетке находится ближе к анимальному полюсу в силу не совсем равномерного распределения желтка, располагаясь в свободной от желточных включений части клетки. Созревание яйцеклетки происходит в воде. Первое редукционное тельце отделяется на анимальном полюсе ооцита еще до оплодотворения. Оно смывается водой и погибает.
Самки ланцетника выметывают яйца в воду, сюда же самцы выпускают сперматозоиды - оплодотворение наружное, моноспермное. После проникновения сперматозоида вокруг яйцеклетки образуется оболочка оплодотворения, которая препятствует проникновению в яйцо других
избыточных спермиев. Вслед за этим происходит отделение второго редукционного тельца, которое располагается между желточной оболочкой и яйцеклеткой.
Все дальнейшее развитие проходит также в воде. Через 4-5 дней из яйцевой оболочки вылупливается микроскопическая личинка, которая переходит к самостоятельному питанию. Сначала она плавает, а затем оседает на дно, растет и совершает метаморфоз
ДРОБЛЕНИЕ. БЛАСТУЛА
Малое количество желтка объясняет простоту дробления и гаструляции. Дробление полное, почти равномерное, радиального типа, в результате образуется целобластула (рис. 1).
Рис. 1. Дробление яйца ланцетника (по Алмазову, Сутулову, 1978):
А - зигота; Б, В, Г - образование бластомеров (показано расположение веретена деления)
Анимальный полюс приблизительно соответствует будущему переднему концу тела личинки. Оплодотворенная яйцеклетка (зигота) целиком дробится на бластомеры в правильной геометрической прогрессии. Бластомеры почти одинаковой величины, анимальные лишь не-
сколько мельче вегетативных. Первая борозда дробления - меридиональная, проходит через анимальный и вегетативный полюс. Она разделяет шаровидное яйцо на две совершенно симметричные половины, но бластомеры округляются. Они шаровидные, имеют малую площадь со-
прикосновения. Вторая борозда дробления также меридиональная, перпендикулярна первой, а третья - широтная.
По мере увеличения количества бластомеров они все больше расходятся от центра зародыша, образуя посередине большую полость. В конце концов, зародыш принимает форму типичной целобластулы - пузырька со стенкой, образованной одним слоем клеток - бластодермой и с полостью, заполненной жидкостью - бластоцелем (рис. 2).
Клетки бластулы, вначале округлые и потому не плотно сомкнутые, затем приобретают форму призм и плотно смыкаются. Поэтому позднюю бластулу, в противоположность ранней, называют эпителиальной.
Стадия поздней бластулы завершает период дробления. К концу этого периода размеры клеток достигают минимума, а общая масса зародыша не увеличивается по сравнению с массой оплодотворенной яйцеклетки.


Рис. 2. Бластула ланцетника (по Алмазову, Сутулову, 1978):
А - внешний вид; Б - поперечный разрез (стрелка показывает задне-переднее направление тела будущего зародыша); В - расположение материалов будущих органов на сагитальном разрезе бластулы
ГАСТРУЛЯЦИЯ
Гаструляция происходит путем инвагинации - впячивания вегетативного полушария бластулы внутрь, по направлению к анимальному полюсу (рис. 3). Процесс протекает постепенно и заканчивается тем, что все вегетативное полушарие бластулы уходит внутрь и становится внутренним зародышевым листком - первичная энтодерма зародыша. Фактор, обуславливающий инвагинацию - разница темпов деления клеток в краевой зоне и в вегетативной части бластулы, приводящей к активному перемещению клеточного материала. Анимальное полушарие становится
.jpg)
Рис. 3. Начальные стадии гаструляции ланцетника (по Мануиловой, 1973):
наружным зародышевым листком - первичная эктодерма. Зародыш приобретает вид двуслойной чаши с широко зияющим отверстием - первичным ртом или бластопором. Полость, в которую ведет бластопор, называют гастроцель (полость первичного кишечника). Бластоцель в результате впячивания низводится до узкой щели между наружным и внутренним зародышевыми листками. На данной стадии зародыш носит название гаструлы (рис. 4 А, Б).
Первичный кишечник (архентерон), представленный внутренним зародышевым листком, окружающим полость гаструлы, является зачатком не только пищеварительной системы, но и других органов и тканей личинки. Бластула, как и яйцеклетка, плавает анимальным полюсом кверху в силу большего веса вегетативного полушария.
В результате инвагинации центр тяжести зародыша перемещается и гаструла поворачивается бластопором кверху.
Бластопор окружен дорсальной, вентральной и боковыми губами. Далее происходит концентрическое смыкание краев бластопора и удлинение зародыша. У ланцетника, представителя вторичноротых, бластопор соответствует не ротовому, а заднепроходному отверстию, обозначая
задний конец зародыша. В результате смыкания краев бластопора и выпячивания тела в передне-заднем направлении, зародыш удлиняется. При этом поперечник гаструлы уменьшается - общая масса составляющих зародыш клеток не может увеличиваться, пока развитие идет под покровом яйцевых оболочек. Зародыш приобретает билатеральную симметрию.
Расположение зачатков в поздней гаструле лучше всего видно на поперечном разрезе зародыша (рис. 4 В, Г).
Наружную стенку его образует эктодерма, неоднородная в своем составе. В дорcальной части эктодерма утолщена и состоит из высоких цилиндрических клеток. Это зачаток нервной системы, которая остается
Рис. 4.


Гаструла ланцетника (по Мануиловой, 1973):
А - ранняя стадия; Б - поздняя стадия; В - поперечный разрез через позднюю гаструлу; Г - гаструла, переходящая в нейрулу (поперечный разрез)
еще на поверхности и образует так называемую медуллярную или нервную пластинку. Остальная эктодерма состоит из мелких клеток и является зачатком покрова животного. Под нервной пластинкой во внутреннем зародышевом листке располагается зачаток хорды, по обеим сторонам которого в виде двух тяжей находится материал мезодермы. В брюшной части располагается энтодерма, образующая основание первичной кишки, крышу которой составляют зачатки хорды и мезодермы .
Материал будущих внутренних органов, находясь в бластуле снаружи, в процессе гаструляции перемещается внутрь зародыша и располагается на местах, развивающихся из них органов. Только зачаток нервной системы остается еще на поверхности. Он погружается внутрь зародыша на стадии, следующей за гаструлой.
НЕЙРУЛЯЦИЯ И ОБРАЗОВАНИЕ ОСЕВЫХ ОРГАНОВ
По окончании гаструляции начинается следующий этап в развитии зародыша - дифференцировка зародышевых листков и закладка органов. Наличие комплекса спинных органов: нервной трубки, хорды и осевой мускулатуры, известных также под именем осевых, является одной их
характерных черт типа хордовых.
Стадия, на которой происходит закладка осевых органов, называется нейрулой. Внешне она характеризуется изменениями, происходящими с зачатком нервной системы.
Они начинаются с нарастания эктодермы по краям нервной пластинки. Образующиеся нервные валики растут навстречу друг другу и затем смыкаются. Пластинка же погружается внутрь и сильно прогибается (рис. 5).

Рис. 5.
Нейрула ланцетника (по Мануиловой, 1973):
А - ранняя стадия (поперечный разрез); Б - поздняя стадия (поперечный
разрез), буквой “ Ц ” обозначена вторичная полость тела (целом)
Это приводит к образованию желобка, а затем нервной трубки, которая в передней и задней части зародыша некоторое время остается открытой (указанные изменения удобнее всего проследить на поперечномразрезе зародыша). Вскоре, в задней части тела эктодерма нарастает на бластопор и отверстие нервной трубки, закрывая их таким образом, что нервная трубка остается сообщенной с кишечной полостью - образуется нервно-кишечный канал.
Одновременно с формированием нервной трубки существенные изменения происходят и во внутреннем зародышевом листке. Из него постепенно обособляются материалы будущих внутренних органов. Зачаток хорды начинает выгибаться, выделяется из общей пластинки и превращается в обособленный тяж в виде сплошного цилиндра. Одновременно происходит обособление мезодермы. Этот процесс начинается споявления небольших карманообразных выростов по двум сторонам
внутреннего листка. По мере роста они отделяются от энтодермы и в ви-де двух тяжей с полостью внутри располагаются по всей длине зародыша. Кроме продольных желобков от переднего конца первичного кишечника последовательно отчленяются еще две пары целомических мешков.
Таким образом, в развитии ланцетника имеется стадия, характеризующаяся наличием трех пар сегментов и свидетельствующая об эволюционном родстве ланцетника с трехсегментарными личинками полухордовых и иглокожих. У ланцетника ярко выражен энтероцельный способ
образования целома - его отшнуровка от первичного кишечника. Этот способ является исходным для всех вторичноротых животных, но почти ни у кого из вышестоящих позвоночных, за исключением круглоротых, с такой ясностью не представлен. После отделения хорды и мезодермы
края энтодермы постепенно сближаются в спинной части и в конце концов смыкаются, образуя замкнутую кишечную трубку.
В ходе дальнейшего развития мезодерма сегментируется: тяжи разделяются поперечно на первичные сегменты или сомиты. Из них образуются три основные закладки:
Дерматом формируется из наружной, обращенной к эктодерместенке сомита, - из его клеток впоследствие возникает соединительная часть кожи, представленная преимущественно фибробластами;
Склеротом образуется из внутренней части сомита, примыкающейк хорде (низшие позвоночные) или к хорде и нервной трубке (высшие позвоночные) - представляет зачаток осевого скелета;
Миотом представляет часть сомита, расположенную между дерматомом и склеротомом - является зачатком всей поперечно-полосатой мускулатуры.
Дифференцировка сомитов у ланцетника протекает иначе, чем у позвоночных. Это различие выражается в том, что у позвоночных сегментируется только спинная часть мезодермальных тяжей, тогда как у ланцетника они полностью распадаются на сегменты. Последние вскоре разделяются на спинную часть - сомиты, и брюшную - спланхнотом.
Сомиты, из которых развивается туловищная мускулатура, остаются обособленными друг от друга, спланхнотомы же сливаются на каждойстороне, образуя левую и правую полости, которые затем объединяются под кишечной трубкой в общую вторичную полость тела (целом).
В развитии ланцетника, с одной стороны, ясно представлены черты типичных позвоночных (характерное расположение зачатков при гаструляции, формирование хорды из дорсальной стенки первичной кишки и нервной пластинки из дорсальной эктодермы), а с другой - черты беспозвоночных вторичноротых животных (целобластула, инвагинационная гаструла, трехсегментная стадия, энтероцельная закладка мезодермы и образования целома).
В дальнейшем, в связи с образованием хвоста нервно-кишечный канал исчезает. В головной части кишечной трубки прорывается ротовое отверстие, а на заднем конце, под хвостом, образуется анальное - путем вторичного прорыва стенки тела животного на месте закрывшегося бластопора. Зародыш переходит в стадию свободноплавающей личинки.
Характерные особенности биологии и анатомии Acrania Бесчерепные – немногочисленная группа морских бентосных животных, типичным представителем которых является ланцетник (Branchiostoma lanceolatum, ранее называемый Amphioxus lanceolatum) Бесчерепные – немногочисленная группа морских бентосных животных, типичным представителем которых является ланцетник (Branchiostoma lanceolatum, ранее называемый Amphioxus lanceolatum) А А

Передний отдел тела Branchiostoma lanceolatum Передняя часть тела ланцетника (внешний вид – слева и схема – справа). На левом снимке видно обращённое вперёд и вниз предротовое отверстие, обрамлённое бордюром из щупалец. Миомеры в виде шевронов составляют мускулатуру от самого переднего конца до самой задней точки тела. Видны первые две камеры сегментированного яичника. На схеме (слева) показана достигающая самой передней точки тела хорда и перфорированная многочисленными жаберными щелями глотка.

Оплодотворённая яйцеклетка Branchiostoma belcheri Фото выполнены с помощью сканирующего электронного микроскопа. Стрелками обозначено второе полярное тельце. На правом фото при боль- шем увеличении на поверхности зиготы можно видеть полярное тельце и многочисленные микроворсинки


Стадия 8 бластомеров – результат третьего «широтного» деления (левый сни- мок - вид с анимального полюса, правый – сбоку, под углом). На фото отчётливо видна разница в размерах между анимальными и вегетативными бластомерами. По-прежнему, невелика поверхность прилегания бластомеров друг к другу.

Branchiostoma belcheri. Стадия 16 бластомеров. При наблюдении с анимально- го полюса (левый снимок) хорошо видны различия в размерах между анималь- ными и вегетативными бластомерами. Следует отметить строгую соосность между бластомерами анимального и вегетативного яруса.







Branchiostoma belcheri. Стадия поздней гаструлы (8 ч. 50 мин.) после оплодо- творения. Типичный двухслойный зародыш. На фото слева можно видеть вы- раженное сужение бластопора (Bp). На сколе справа заметны различия между эктодермой (слой кубических клеток) и эндодермой (слой цилиндрических кле- ток). Ac – архентерон (гастроцель).


 19
Нотогенез у ланцетника Формирование осевого комплекса зачатков (поперечные срезы). Выгибание нейральной пластинки и хордальной пластинки в противоположных направлениях. Выпячивание и отделение мезодермальных валиков (7) из свода архентерона с последующей их сегментацией.
19
Нотогенез у ланцетника Формирование осевого комплекса зачатков (поперечные срезы). Выгибание нейральной пластинки и хордальной пластинки в противоположных направлениях. Выпячивание и отделение мезодермальных валиков (7) из свода архентерона с последующей их сегментацией.

Предличиночные стадии развития. Происходит смыкание нейральных вали- ков (6), обособление хорды (5), образование сомитов и затем целомических мешков (9). 10 – миотом, 11 – миоцель, 12 – дерматом, 13 – закладка вен- трального мезентерия, 14 – подкишечная вена (черного цвета), 15 – висце- ральный листок спланхнотома, 16- париетальный листок спланхнотома, 17 – спланхнотом (латеральная пластинка мезодермы), 18 – спланхноцель.


Гаструляция у ланцетника представляет удобную модель для изучения общих закономерностей этого процесса у типа хордовых. Этот процесс, как было показано ранее, у ланцетника, происходит по типу инвагинации. На ранней стадии гаструлы обнаруживается два зародышевых листка: эктодермальный, образованный из крыши бластулы, и энтодермальный- из дна бластулы. Гастроцель сообщается с внешней средой при помощи бластопора.
Наступает следующий этап развития зародыша - образование третьего зародышевого листка-мезодермы. Клетки дорзальной губы бластопора, быстро размножаясь, погружаются в составе энтодермы и на дорзальной ее стороне образуют хордальную пластинку- зачаток будущей хорды. Мелкие клетки вентральной и боковых губ бластопора также врастают в энтодермальный листок и располагаются в его составе по сторонам от хордальной пластинки.
Двуслойный зародыш приобретает вытянутую форму, имея в своем составе материал трех зародышевых листков. Материал мезодермы занимает дорзальную часть внутреннего листка. Из первичной эктодермы, расположенной над хордальной пластинкой, выделяется нервная пластинка. Последняя погружается и по длине зародыша образует нервный желобок. Нервный желобок, замыкаясь кверху, образует нервную трубку.
Хордальная пластинка выходит из состава первичного внутреннего листка, замыкается книзу и образует плотный клеточный тяж - хорду.
Одновременно мелкоклеточный материал, находившийся в первичном внутреннем листке по сторонам от хорды, выпячивается в дорзальном направлении в виде двух карманов. Вначале эти карманы открываются в гастроцель, а затем отделяются в виде двух замкнутых складок, расположенных вдоль гаструлы.
Так образуется третий зародышевый листок - мезодерма. После выделения хорды и мезодермы материал первичного внутренного листка смыкается, образуя энтодерму - внутренний зародышевый листок (рис. 5).
ГАСТРУЛЯЦИЯ у птиц
Гаструляция у птиц представляет довольно близкую модель процессам, имеющим место у высших хордовых, в том числе и у человека.
У птиц в результате дискоидального (меробластического) дробления образуется бластодиск. Он состоит из клеток неправильной формы, плотно прилегающих друг к другу и расположенных на огромной массе нераздробившегося желтка. Под влиянием ферментов клеток бластодиска часть желтка разжижается и образуется полость, заполненная жидкостью - бластоцель. Крыша этой бластулы представлена бластодиском, дно - нераздробившейся массой желтка. Бластоцель - подзародышевая полость.
Затем наступает гаструляция, состоящая из двух фаз. Первая фаза начинается до откладывания яйца и заключается в обособлении энтодермы путем деляминации клеток бластодиска. В результате возникает двуслойный зародыш. Верхний слой клеток дискобластулы приобретает призматическую форму и располагается в один правильный ряд. Нижний слой клеток дискобластулы сохраняет округлую или неправильную форму и лежит беспорядочно на желтке. Между клетками верхнего и нижнего слоя возникает полость - гастроцель.
Вторая фаза гаструляции начинается во второй половине первых суток инкубации. Бластодиск разрастается, в центре его выделяется зародышевый щиток, из которого в дальнейшем развивается тело зародыша. Окружающая зародышевый щиток часть дискобластулы представляет внезародышевый материал, в котором различают светлое и темное поле. Светлое поле расположено вокруг зародышевого щитка и состоит из клеток, отделившихся от желтка подзародышевой полостью в силу частичного использования желтка зародышем. Темное поле занимает периферию дискобластулы и состоит из клеток, плотно прилегающих к желтку и растущих по его поверхности. К этому моменту резко выражено разделение клеточного материала на зародышевую (зародышевый щиток) и внезародышевую (светлое и темное поле) часть.
В течение 24 часов насиживания или инкубации на зародышевом щитке в результате размножения клеток наблюдается перемещение их спереди назад по периферии зародышевого щитка. Оба потока клеток встречаются в центре у заднего конца зародышевого щитка, сливаются и перемещаются кпереди посредине зародышевого щитка. В результате образуется утолщенный тяж клеток, получивший название первичной полоски.
На переднем конце первичной полоски формируется утолщение- первичный узелок. В дальнейшем часть бластомеров, расположенных кпереди от первичного узелка, перемещается к последнему, подворачивается под эктодерму и формирует растущий кпереди между эктодермой и энтодермой головной или хордальный вырост - зачаток хорды.
Бластомеры задней половины зародышевого щитка смещаются к первичной полоске и через эту область погружаются под эктодерму, располагаясь в полости между эктодермой и энтодермой. Этот материал образует зачаток мезодермы - третьего зародышевого листка. Клетки хордального выроста размножаются и. перемещаются, занимая центральное положение между эктодермой

Рис.%6. Развитие комплекса осевых органов у птиц 1-эктодерма; 2 -нервный желобок; 3 - нервная трубка; 4 - амнион; 5 - сомиты; 6 - париетальный листок сплахнотома; 7 - висцеральный листок спланхнотома; 8 - хорда; 9 - дерматом; 10 - миотом; 11 - склеротом; 12 - нефротом; 13 - аорта; 14 - энтодерма; 15 - амниотическая складка и энтодермой в передней половине зародышевого щитка Вслед за этим клетки мезодермы также перемещаются в переднюю половину зародышевого щитка, располагаясь между эктодермой и энтодермой по бокам от хордального выроста. Это парахордальная мезодерма. Ее можно подразделить на дорзальную и вентральную.
По мере того как клетки первичной полоски уходят в закладки хорды и мезодермы на поверхности остается материал эктодермы и входящей в нее нервной пластинки. Таким образом, первичная полоска и головной узелок по своему значению соответствуют бластопору ланцетника, т. к. в этом месте проходит перемещение зародышевого материала, приводящее к образованию 3-зародышевых листков. Наиболее глубоко расположен зачаток кишечной энтодермы. Он непосредственно прилегает к поверхности желтка и находится во внутреннем слое дискобластулы.
К концу гаструляции в теле зародыша имеются все зачатки, из которых возникает типичный для хордовых комплекс осевых органов (рис. 6.).
Результат активного деления клеток, роста и направленных перемещений (миграций) клеточных потоков с формированием многослойного зародыша, или гаструлы, (возникновением послойно расположенных, отделенных друг от друга отчетливой щелью, зародышевых листков: наружного - эктодермы, среднего - мезодермы, внутреннего - энтодермы). Перемещение клеток происходит в строго определенной области зародыша - в области серого серпа. Последний был описан В. Ру в 1888 г. В оплодотворенном яйце амфибии серый серп выявляется как окрашенная область на стороне, противоположной проникновению спермия. В этом месте, как полагают, локализуются факторы, необходимые для гаструляции.
У разных представителей позвоночных совершается несколькими основными способами: путем инвагинации (впячивания), иммиграции (перемещения части клеток внутрь зародыша), эпиболии (обрастания), деламинации (расщепления). Способы гаструляции зависят от типа яйцеклетки. При любом способе гаструляции в качестве ведущих сил выступают неравномерная пролиферация клеток в разных частях зародыша, уровень обменных процессов в клетках, расположенных в разных частях зародыша, активность амебоидных движений клеток, а также индуктивные факторы (белки, нуклеопротеиды, стероиды и др.). В результате гаструляции обособляются основные зачатки органов и тканей.
Следующим периодом эмбриогенеза является гисто- и органогенез - дифференцировка различных тканей и органов организма из материала зародышевых листков и эмбриональных зачатков.
В результате гаструляции возникает многослойный зародыш . Несмотря на различные способы гаструляции после выделения материала зародышевых листков по оси зародыша находится материал хорды, который подстилает нервную пластинку, слева и справа от хорды располагается материал мезодермы. Все это характеризует осевой комплекс зачатков. В дальнейшем происходит формирование зачатков органов, представляющих собой пространственно локализованные группы стволовых клеток - источников развития тканей. Закономерности дифференцировки клеточного материала зачатков можно проследить в эмбриогенезе наиболее изученных животных.
Ланцетник. Развитие ланцетника.
Классическим объектом эмбриологических исследований ланцентник , подробно изученный А.О. Ковалевским. Ланцетник является представителем класса хордовых животных подтипа бесчерепных, размером до 8 см и обитает на песчаном дне в теплых морях. Свое название получил из-за формы, напоминающей ланцет (хирургический инструмент с обоюдоострым лезвием, современный скальпель).
Яйцеклетка ланцетника олиго- и изолецитальная, размером 110 мкм, ядро располагается ближе к анимальному полюсу. Оплодотворение наружное. Дробление зиготы полное, почти равномерное, синхронное и заканчивается образованием бластулы. В результате чередования меридианальных и широтных борозд дробления формируется однослойная бластула с полостью, заполненной жидкостью - бластоцелем. Бластула сохраняет полярность, ее дно представляет собой вегетативную, а крыша - анимальную части; между ними находится краевая зона.
При гаструляции происходит впячивание вегетативной части бластулы в анимальную. Впячивание постепенно углубляется и, наконец, формируется двустенная чаша с широко зияющим отверстием, ведущим в новообразованную полость зародыша. Такой способ гаструляции называется инвагинацией. Так бластула превращается в гаструлу. В ней материал зародыша оказывается дифференцированным на наружный листок - эктодерму, и внутренний - энтодерму. Полость чаши именуется гастроцелем, или полостью первичной кишки, которая сообщается с внешней средой посредством бластопора, что соответствует заднепроходному отверстию. В бластопоре различают дорсальную, вентральную и две латеральных губы. В результате инвагинации центр тяжести зародыша смещается, и зародыш поворачивается бластопором кверху. Постепенно происходит смыкание краев бластопора и удлинение зародыша. Топография клеток в составе губ бластопора определяет развитие разных частей зародыша. При гаструляции из внутреннего листка гаструлы обособляются хорда и мезодерма, которые располагаются между экто- и энтодермой. Завершается гаструляция образованием осевого комплекса зачатков и дальше - обособлением зачатков органов. Хорда индуцирует развитие нервной трубки из материала дорсальной эктодермы. Эта часть эктодермы утолщается, образуется нервная пластинка (нейроэктодерма), которая прогибается по средней линии и превращается в желобок.

Края желобка постепенно смыкаются в нервную трубку. Оставшаяся часть эктодермы - кожная, срастается над нервной трубкой. Однако на самом переднем и заднем концах зародыша нервная трубка некоторое время сообщается с внешней средой с помощью двух отверстий - нейропоров. В последующем происходит разделение мезодермы на спинные сегменты - сомиты, количество которых нарастает от 15 пар до 60-65 пар у взрослого ланцетника. Часть латерально расположенной мезодермы не сегментируется и расщепляется на наружный (париетальный) и внутренний (висцеральный) листки спланхнотома. Эти листки растут между экто- и энтодермой и, достигнув середины на вентральной стороне зародыша под кишечной трубкой, срастаются, формируя единую вторичную полость - целом. На переднем конце зародыша возникает углубление (ротовая бухта), растущее по направлению к переднему отделу кишечной трубки. При контакте эктодермы ротовой бухты и слепого конца кишечной трубки происходит апоптоз клеток и возникает сообщение кишки с внешней средой. Аналогичный процесс протекает на заднем конце зародыша. По бокам головного отдела зародыша также возникает контакт кожной эктодермы и кишечной энтодермы. В месте этого контакта происходит прорыв. Так полость передней кишки сообщается с внешней средой (формируется жаберный аппарат). После этого зародыш выходит из яйцевой оболочки во внешнюю среду в виде личинки.
Методы маркирования для изучения процессов миграции бластомеров позволили выделить определенные области зародыша на ранних стадиях развития (зиготы - бластулы), которые позднее развиваются в зародышевые листки и эмбриональные зачатки органов и тканей. Эти области были названы презумптивными (предполагаемыми) участками, или зачатками.