Кроме очень большой кольцевой хромосомы, расположенной в ядерной зоне, большинство бактерий содержат одну или несколько небольших кольцевых молекул ДНК, которые находятся в свободном состоянии в цитоплазме клетки.
Рис. 27-20. Электронные микрофотографии плазмид, выделенных из двух видов бактерий. А. Плазмида из Е. coli, которая придает клеткам устойчивость к тетрациклину. Б. Плазмиды из Neisseria gonorrhoeae - бактерии, вызывающей гонорею. Большинство молекул находится в релаксированном состоянии. На примере скрученных сверхспиралъных плазмид (одна из них в центре фотографии) видно, насколько эффективной может оказаться компактная упаковка колъцбвой ДНК в рефльтате отрицательных поворотов.
Эти экстрахромосомные элементы называют плазмидами (рис. 27-20). В большинстве своем плазмиды очень малы и содержат всего лишь несколько генов, т. е. гораздо меньше, чем бактериальная хромосома, в которой генов тысячи. В некоторых клетках, однако, плазмиды могут быть достаточно большими. Плазмиды несут генетическую информацию и реплицируются с образованием дочерних плазмид, которые при делении клетки переходят в дочерние клетки. В течение многих циклов деления клетки плазмиды «живут» своей собственной, обособленной от хромосомной ДНК «жизнью». Однако иногда они могут встраиваться в хромосомную ДНК и с таким же успехом покидать ее.
Некоторые плазмиды несут гены, которые определяют устойчивость бактерии-хозяина к антибиотикам, например тетрациклину и стрептомицину. Бактерии, содержащие такие плазмиды, устойчивы к этим антибиотикам. При лечении бактериальной инфекции антибиотиком эти клетки могут выжить в организме человека, так как антибиотик убивает только чувствительные к нему клетки. Устойчивые к антибиотику клетки могут затем размножаться и вызывать инфекцию, которую нельзя сдержать антибиотиком.
По этой причине антибиотики не следует применять для лечения инфекций без разбора: всегда необходимо быть уверенным в том, что возбудитель данной болезни чувствителен к используемому антибиотику. Плазмиды могут переноситься из устойчивых к антибиотику клеток в чувствительные клетки того же или другого вида, делая эти клетки устойчивыми.
Другая важная особенность плазмид состоит в том, что их можно очень легко выделить из бактериальных клеток. В выделенную плазмиду можно встроить новые гены из других видов и затем такую модифицированную плазмиду ввести обратно в обычную для нее клетку-хозяина. Такая плазмида, несущая чужеродный ген, будет реплицироваться и транскрибироваться и может заставить клетку-хозяина синтезировать белки, кодируемые искусственно встроенным геном, даже если он не является частью нормального генома клетки. Позже мы увидим, как получают такие рекомбинантные ДНК, как они транскрибируются и транслируются с образованием потенциально полезных продуктов.
ДНК-содержащие вирусы имеют либо собственные ферменты репликации (в капсиде), либо в их геноме закодирована информация о синтезе вирусных ферментов, обеспечивающих репликацию вирусной нуклеиновой кислоты. Количество этих ферментов различно в применении к разным вирусам. Например, в геноме бактериального вируса Т4 закодирована информация о синтезе около 30 вирусных ферментов. Далее геном крупных вирусов кодирует нуклеазы, разрушающие ДНК клетки-хозяина,а также белки, воздействие которых на клеточную РНК-полимеразу сопровождается тем, что «обработанная таким образом РНК-полимераза транскрибирует на разных стадиях вирусной инфекции разные вирусные гены. Напротив, малые по размерам ДНК-содержащие вирусы в большей мере зависят от ферментов клеток-хозяев. Например, синтез ДНК аденовирусов обеспечивается клеточными ферментами.[ ...]
Бактериальные ДНК - это высокополимерные соединения, состоящие из большого числа нуклеотидов - полинуклеотиды с молекулярным весом около 4 млн. Молекула ДНК представляет собой цепь нуклеотидов, где расположение их имеет определенную последовательность. В последовательности расположения азотистых оснований закодирована генетическая информация каждого вида. Нарушение этой последовательности возможно при естественных мутациях или же под влиянием мутагенных факторов. При этом микроорганизм приобретает или утрачивает какое-либо свойство. У него наследственно изменяются признаки, т. е. появляется новая форма микроорганизма. У всех микроорганизмов - прокариотов и эукариотов - носителями генетической информации являются нуклеиновые кислоты - ДНК и РНК. Лишь некоторые вирусы представляют собой исключение: у них ДНК отсутствует, а наследственная информация записана или отражена только в РНК.[ ...]
В бактериальных клетках в общем количестве оснований ДНК 32-65 мол.% гуанина и цитозина.[ ...]
Ядро бактериальной клетки. Примерно 1-2% веса сухой массы микроорганизмов приходится на ДНК, в которой заложена генетическая информация организма. У большинства микроорганизмов имеются области (или несколько областей), в которой сконцентрировано основное количество ДНК, имеющие определенную структуру (или органеллу) и называющиеся ядром. Ядро (или ядерное вещество) связано с цитоплазматической мембраной, независимо от того, окружено оно элементарными мембранами (как у амебы) или не имеет их (как у бактерий и сине-зеленых водорослей). Ядерное вещество активизируется в период размножения и при наступлении возрастных изменений, связанных со старением клетки.[ ...]
Сегмент ДНК (ген), который предназначен для молекулярного клонирования, должен обладать способностью к репликации при переносе его в бактериальную клетку, т. е. быть репликоном. Однако он такой способностью не обладает. Поэтому, чтобы обеспечить перенос и обнаружение клонируемых генов в клетках, их объединяют с так называемыми генетическими векторами. Последние должны обладать, как минимум, двумя свойствами. Во-первых, векторы должны быть способны к репликации в клетках, причем в нескольких копиях. Во-вторых, они должны обеспечивать возможность селекции клеток, содержащих вектор, т. е. обладать маркером, на который можно вести контрселекцию клеток, содержащих вектор вместе с клонируемым геном (рекомбинантные молекулы ДНК). Таким требованиям отвечают плазмиды и фаги. Плазмиды являются хорошими векторами по той причине, что они являются репликонами и могут содержать гены резистентности к,какому-либо антибиотику, что позволяет вести селекцию бактерий на устойчивость к этому антибиотику и, следовательно, легкое обнаружение рекомбинантных молекул ДНК.[ ...]
У бактерий ДНК упакована менее плотно, в отличие от истинных ядер; нуклеоид не обладает мембраной, ядрышком и набором хромосом. Бактериальная ДНК не связана с основными белками - гистонами - ив нуклео-иде расположена в виде пучка фибрилл.[ ...]
Применение методов рекомбинантной ДНК для получения биологических агентов для борьбы с загрязнениями находится на ранней стадии, но есть метод, который может оказаться полезным в обозримом будущем - это генетическое зондирование. Отбор организмов, способных трансформировать новое соединение, часто основан на способности использовать вещество как субстрат роста. Если рост слабый или субстрат только комета-болизируется, то методы селекции окажутся непригодными для идентификации деградативной способности. Следовательно, было бы полезно развивать генетическое зондирование для определения специфических последовательностей в плазмидах и хромосомах, это необходимо для определения катаболического потенциала, даже если этот потенциал не экспрессируется. Такие зонды разработаны для TOL-плазмид. Метод может определить одну бактериальную колонию, содержащую TOL-плазмиду, среди 106 колоний Escherichia coli. Такой мощный инструмент будет иметь огромное значение для выделения скрытых катаболических функций .[ ...]
Разработка изящной методики «клонирования» ДНК для получения большого количества точных копий специфических фрагментов ДНК (рис. 13.4) открыла в последнее время новые горизонты в изучении структуры, организации и функции генома. Если расщепить двухцепочечную ДИК одним из ферментов «рестрикции» (одной из нуклеаз), специфично узнающих и расщепляющих короткие последовательности нуклеотидов (4- 6 пар), то возникают в высшей степени воспроизводимые фрагменты ДНК. Концы двух цепей ДНК обычно бывают смещены относительно друг друга вследствие специфичности мест разрезания двухцепочечной молекулы, цепи которой комплементарны по составу оснований. ДНК обычно встраивают в плазмидный ген, важный для селекции, такой, как ген устойчивости к антибиотикам, что позволяет содержащим такую плазмиду бактериям расти в присутствии антибиотика.[ ...]
В бактериях при репликации образуется много копий плазмид, и таким образом можно «вырастить» большие количества встроенных фрагментов ДНК, а затем снова просто выделить их путем расщепления тем же самым ферментом рестрикции с разделением полученных продуктов гель-электрофорезом. Использование этого метода рекомбинации ДНК произвело революцию в изучении генов.[ ...]
Недавно было обнаружено, что мутагенное действие на бактериальные-ДНК-вирусы оказывают лучи с длиной волны 320-400 нм (область, близкая к зоне видимого света), обладающие низкой интенсивностью . Возможное влияние радиации в этом диапазоне длин волн на вирусы растений пока не обнаружено.[ ...]
Кривые зависимости реассоциацни от СОТ, полученные для бактериальной ДНК, лишены перегибов, а ДИК эукариот реас-социирует по другому типу (рис. 13.2). При низких концентрациях ДНК и коротком времени инкубации реиатурирует заметная доля одноцепочечной ДНК, а при увеличении СОТ образуется дополнительное количество двухцепочечных молекул, так что получается двухфазная кривая. Быстрая реиатурация при низких значениях СОТ показывает, что какие-то последовательности у эукариот повторяются много раз, т. е. до 10 000 раз и более.[ ...]
Отсутствие СХС может иммитироваться и в тех случаях, когда ДНК тестерных фагов не содержат сайтов, узнаваемых существующей в исследуемом штамме рестриктазой. Это явление представляет собой один из вариантов эволюционных адаптивных изменений бактериальных вирусов, призванных способствовать преодолению ими барьера СХС. Действие давления отбора в данном конкретном случае выражается в статистическом достоверном уменьшении числа или даже полной элиминации в фаговой ДНК последовательностей нуклеотидов, являющихся субстратом рестриктаз, характерных для клеток-хозяев бактериального вируса .[ ...]
Линдегрен описал возможные стадии образования бактериофага из ДНК профага, предположив, что профаг возникает как фрагмент чужеродной бактериальной ДНК, случайно проникший в клетку, который на ранних стадиях делится синхронно с бактериальной ДНК. Следующим важным этапом в развитии вируса явилось бы такое изменение профага, в результате которого стало возможным его независимое от ДНК клетки-хозяина размножение; в результате профаг использовал бы. все доступные нуклеотиды, нарушив тем-самым рост клетки-хозяина. Наконец, на какой-то более поздней стадии могла бы образоваться защитная белковая оболочка и возникли другие белки, что должно было обеспечить выживание ДНК вне организма хозяина и эффективное заражение новых клеток. Отделившийся фрагмент бактериальной ДНК вначале, очевидно, кодировал белки, приспособленные к бактериальным функциям. Необходимы очень существенные изменения в ДНК, чтобы могли возникнуть объекты настолько сложные и специализированные, как, скажем, фаг Т2 Е. coli, содержащие к тому же основания, которые в бактериальной ДИК отсутствуют.[ ...]
Генетическая информация бактерий не ограничивается ДНК, расположенной в нуклеоиде бактериальной клетки. Как уже отмечалось в предыдущих разделах книги, носителями наследственных свойств служат также внехромосомные элементы, получившие общее название плазмид. В отличие от ДНК ядерных эквивалентов-нуклеоидов, являющихся органоидами бактериальной клетки, плазмиды представляют собой независимые генетические элементы. Потеря плазмид или их приобретение не отражается на биологии клетки (приобретение плазмид оказывает положительное влияние лишь на популяцию в целом, повышая жизнеспособность вида). К трансмиссивным относят плазмиды, инициирующие свойства доноров у клеток-хозяев. При этом последние получают новое качество - возможность конъюгировать с клетками-реципиентами и отдавать им свои плазмиды. Клетки-реципиенты, приобретая во время конъюгации плазмиды, сами превращаются в доноров.[ ...]
Отсутствие адсорбции не исчерпывает разнообразия вариантов взаимодействия бактериальных вирусов и микробных клеток. Они иллюстрируют лишь одну сторону этого явления, а именно проявление клеточных защитных механизмов, фенотипически (по критерию отсутствия роста) иммитирующих рестрикцию. Однако, существует и другой вариант взаимодействия клетка-бактериофаг, который может иммитировать отсутствие СХС. Примерами таких механизмов является синтез ингибиторов и метилаз кодируемых фаговыми генами, защищающих вирусную ДНК от действия рестриктаз II типа.[ ...]
Механизм обеззараживающего действия хлора связан с нарушением обмена веществ бактериальной клетки в процессе дезинфекции воды. При этом выявлено влияние на ферментную активность бактерий, в частности, на дегидрогеназы, катализирующие окислительно-восстановительные реакции в бактериальной клетке. А. М. Ски-дальской (1969) было изучено влияние хлора на процесс декарбоксилирования аминокислот бактерий, протекающий в присутствии строго специфичных ферментов-декарбоксилаз, а также был определен нуклеотидный состав ДНК кишечной палочки после окончания процесса обеззараживания при различных уровнях бактерицидного эффекта.[ ...]
Бактериофаги Т-группы имеют форму барабанных палочек размером 100 х 25 нм. Их геном представлен ДНК. Они являются вирулентными фагами, т. к. после инфицирования ими бактериальных клеток последние лизируются с освобождением большого количества вновь синтезированных фаговых частиц.[ ...]
Плазмиды бактерий - это генетические структуры, находящиеся в цитоплазме и представляющие собой молекулы ДНК размером от 2250 до 400 ООО пар азотистых оснований. Они существуют обособленно от хромосом в количестве от одной до нескольких десятков копий на одну бактериальную клетку.[ ...]
Штамм Pseu.dom.onas вуг1 ае ри. рка8еоИсо1а обладает плазмидой длиной 150 тыс. п. н., которая может реплицироваться автономно, а может интегрироваться в бактериальную хромосому. Последующее неточное вырезание позволило получить семейство плазмид длиной от 35 до 270 тыс. п. н., некоторые из них содержали большие сегменты хромосомной ДНК .[ ...]
В ходе эволюции бактерии развили способность синтезировать так называемые рестрицирующие ферменты (эндонуклеазы), которые стали частью клеточной (бактериальной) системы рестрикции-модификации. У бактерий системы рестрикции-модификации являются внутриклеточной иммунной системой защиты от чужеродной ДНК. В отличие от высших организмов, у которых распознание и разрушение вирусов, бактерий и других патогенов происходит внеклеточно, у бактерий защита от чужеродной ДНК (ДНК растений и животных, в организме которых они обитают) происходит внухриклеточно, т. е. тогда, когда чужеродная ДНК проникает в цитоплазму бактерий. С целью защиты бактерии в ходе эволюции развили также способность «метить» собственную ДНК метилирующими основаниями на определенных последовательностях. По этой причине чужеродная ДНК из-за отсутствия в ней метальных групп на тех же последовательностях плавится (разрезается) на фрагменты разными бактериальными рестриктазами, а затем деградируется бактериальными экзонуклеазами до нуклеотидов. Можно сказать, что таким образом бактерии защищают себя от ДНК растений и животных, в организме которых они обитают временно (как патогены) или постоянно (как сапрофиты).[ ...]
Наследственные свойства бактерий или отдельные признаки закодированы в единицах наследственности - генах, линейно расположенных в хромосоме вдоль нити ДНК. Следовательно, ген является фрагментом нити ДНК- Каждому признаку соответствует определенный ген, а часто еще меньший отрезок ДНК - кодон. Иначе говоря, в нити ДНК в линейном порядке расположена информация обо всех свойствах бактерий. При этом у бактерий есть еще одна особенность. В ядрах эукариотов содержится обычно несколько хромосом, число их в ядре постоянно у каждого вида. Нуклеоид бактерий содержит лишь одно кольцо из нити ДНК, т. е. одну хромосому. Однако запасом информации, заключенным в одной хромосоме или в кольцеобразно сомкнувшейся двунитчатой спирали ДНК, сумма наследственных признаков бактериальной клетки не исчерпывается. Плазмиды содержат ДНК, также несущую генетическую информацию, передаваемую от материнской клетки к дочерней.[ ...]
Мутации - это изменения в генном аппарате клетки, которые сопровождаются изменениями контролируемых этими генами признаков. Различают макро- и микроповреждения ДНК, ведущие к изменению свойств клетки. Макроизменения, а именно: выпадение участка ДНК (деления), перемещение отдельного участка (транслокация) или поворот определенного участка молекулы на 180° (инверсия) -у бактерий наблюдаются сравнительно редко Гораздо более характерны для них микроповреждения, или точечные мутации, т. е. качественные изменения в отдельных генах, например замена пары азотистых оснований. Мутации бывают прямые и обратные, или реверсивные. Прямые - это мутации организмов дикого типа, например утрата способности самостоятельно синтезировать факторы роста, т. е. переход от прото- к ауксотрофности. Обратные мутации представляют собой возвращение, или реверсию, к дикому типу. Способность к реверсии характерна для точечных мутаций. В результате мутаций изменяются такие важнейшие признаки, как способность самостоятельно синтезировать аминокислоты и витамины (ауксотрофные мутанты), способность к образованию ферментов. Эти мутации называют биохимическими. Хорошо известны также мутации, ведущие к изменению чувствительности к антибиотикам и другим антимикробным веществам. По происхождению мутации разделяют на спонтанные и индуцированные. Спонтанные возникают самопроизвольно без вмешательства человека и носят случайный характер. Частота таких мутаций очень низка и составляет от 1 X Ю“4 ло 1 X 10-10. Индуцированные возникают при воздействии на микроорганизмы физических или химических мутагенных факторов. К физическим факторам, обладающим мутагенным действием, относятся ультрафиолетовое и ионизирующие излучения, а также температура. Химическими мутагенами являются ряд соединений и среди них наиболее активны так называемые супермутагены. В природных условиях и эксперименте изменения в составе бактериальных популяций могут возникать в результате действия двух факторов - мутаций и автоселекции, происходящей в результате адаптации некоторых мутантов к условиям среды обитания. Такой процесс, очевидно, наблюдается в среде, где преобладающим источником питания является синтетическое вещество, например, ПАВ или капролактам.[ ...]
Одиночная клетка E. coli окружена трехслойной клеточной оболочкой толщиной порядка 40 нм, представляющей собой «мешок» или «конверт», в котором заключено клеточное содержимое в виде, примерно, 2 х Ю 1Я г белка, 6 х 10 16 г ДНК и 2 х 10 14 г РНК (в основном ри-босомной РНК). В бактериальной клетке синтезируется около 2000 разных белков, большинство которых содержится в цитоплазме. Концентрация одних белков составляет 10“® М, тогда как других - порядка 2 х 10"4 М (от 10 до 200 000 молекул на клетку).[ ...]
У одноклеточных организмов половое размножение существует в нескольких формах. Конъюгация встречается также у инфузорий, у которых во время этого процесса происходит переход ядер от одних особей к другим, после чего следует деление последних.[ ...]
Бактерии: прокариоты («доядерные») одноклеточные организмы. Их клетки не имеют отделенного от цитоплазмы ядра. Однако генетическая программа, как и у всех живых организмов, закодирована в виде последовательности нуклеотидов в ДНК и несет информацию о структуре белков. Бактериальные клетки не содержат таких органелл, как хлоропласты (специализированные для фотосинтеза) и митохондрии (специализированные для клеточного дыхания и синтеза АТФ). Эти биохимические процессы происходят у бактерий в цитоплазме.[ ...]
Крайне малые размеры клеток являются характерной, но не главной особенностью бактерий. Все бактерии представлены особым типом клеток, лишенных истинного ядра, окруженного ядерной мембраной. Аналогом ядра у бактерий является нуклеоид - ДНК-содержащая плазма, не отграниченная от цитоплазмы мембраной. Кроме того, для бактериальных клеток характерны отсутствие митохондрий, хлоропла-стов, а также особое строение и состав мембранных структур и клеточных стенок. Организмы, в клетках которых отсутствует истинное ядро, называются прокариотами (доядер-ными) или протоцитами (т. е. организмами с примитивной организацией клеток).[ ...]
Микоплазменные клетки имеют овальную форму, а их размеры составляют около 0,1-0,25 нм в диаметре (рис. 43). Для них характерно наличие тонкой наружной плазматической мембраны (толщина - около 8 нм), которая окружает цитоплазму, содержащую молекулу ДНК, достаточную для кодирования около 800 разных белков, РНК разных типов, рибосом диаметром порядка 20 нм. В их цитоплазме содержатся различные включения в виде белков, гранул липидов и других соединений. Из-за недостаточной жесткости клеточкой мембраны микоплазмы проходят через бактериальные фильтры.[ ...]
Установлено, что на рибосомах происходят связывание активированных аминокислот и укладка их в полипептидную цепь в соответствии с генетической информацией, полученной из ядра через информационную (матричную) РНК (мРНК), которая как бы считывает соответствующую информацию с ДНК и передает ее на рибосомы. Целый ряд белков синтезирован на изолированных рибосомах и при этом отмечено включение в них меченых аминокислот. Роль матрицы в белковом синтезе выполняет мРНК, которая прикрепляется к рибосоме. На поверхности последней происходит взаимодействие между комплексом аминокислот, транспортной РНК, несущей очередную аминокислоту, и нуклеотидной последовательностью информационной РНК, которая функционирует на рибосоме однократно и после синтеза полипептидной цепи распадается, а вновь синтезированный белок накапливается в рибосомах. В бактериальной клетке при периоде регенерации 90 мин скорость кругооборота мРНК достигает 4-6 с.[ ...]
Цитоплазма представляет собой коллоидный раствор, дисперсной фазой которого являются сложные белковые соединения и вещества, близкие к жирам, а дисперсионной средой - вода. У некоторых форм бактерий в цитоплазме содержатся включения - капельки жира, серы, гликогена и др. Постоянными составляющими бактериальных клеток являются особые выросты цитоплазматической мембраны - мезосомы, в которых содержатся ферментные окислительно-восстановительные системы. В этих образованиях идут в основном процессы, связанные с дыханием бактерий. В мелких включениях - рибосомах, содержащих рибонуклеиновую кислоту, осуществляется биосинтез белка. Большинство видов бактерий не имеет обособленного ядра. Ядерное вещество, представленное ДНК, у них не отделено от цитоплазмы и образует нуклеоид. Транспортировка веществ, необходимых для жизнедеятельности клетки, и отвод продуктов обмена осуществляется по особым каналам и полостям, отделенным от цитоплазмы мембраной, имеющей такое же строение, как и цитоплазматическая. Это структурное образование называется эндоплазматической сетью (ретикулум).[ ...]
Представление об изменчивости и наследственности бактерий нельзя составить без знания некоторых положений молекулярной генетики прокариотической клетки. В основе процессов приспособления микробных культур к изменяющимся экологическим условиям лежат изменчивость и наследственность, являющиеся разделами генетики бактерий. При изложении цитологии бактериальной клетки уже рассматривалась структура ДНК и РНК и их роль в жизни клетки. Характерное строение ДНК сохраняется у каждого вида и передается потомству из поколения в поколение, как и другие признаки. ДНК бактерий представляет собой двунитчатую спираль, замыкающуюся в кольцо. Кольчатая нить ДНК бактерий, расположенная в ну-клеоиде, не содержит белка. Такое кольцо ДНК соответствует хромосоме эукариотической клетки. Известно, что в хромосоме эукариотических клеток, кроме ДНК, всегда содержится белковый компонент. Отсюда следует, что понятие хромосомы у эукариотов несколько отлично от понятия хромосомы бактерий. Нить ДНК, представляющая собой хромосому бактерий, разумеется, у разных видов различается. Сахарофосфатный компонент ДНК У всех видов бактерий одинаков; расположение азотистых оснований и их комбинация, напротив, различаются у разных видов.[ ...]
Все возрастающее беспорядочное применение антибиотиков в животноводстве, которые используются в малых дозах как стимуляторы роста, а также в качестве превентивной меры против вызванных стрессом желудочно-кишечных расстройств у животных на фермах, приводит к все более широкому распространению в микробных популяциях R-фактора устойчивости к антибиотикам, передающегося от одной бактериальной клетки к другой при конъюгации. Передача происходит через плазмиду, которая представляет собой кольцевую экстрахромосомную ДНК, способную к репликации.[ ...]
В противоположность вирулентным фагам, известны так называемые фаги умеренного действия, или просто умеренные фаги. Типичным представителем таких фагов является фаг X, который тоже использовался и используется в качестве экспериментальной модели для выяснения многих вопросов молекулярной генетики. Фагу X присущи два важных свойства. Подобно вирулентным фагам он может инфицировать бактериальные клетки, размножаться вегетативно, продуцируя в клетках сотни копий и лизировать клетки С освобождением зрелых фагочастиц. Однако ДНК этого фага может включаться в бактериальную хромосому, превращаясь в профаг. При этом происходит так называемая лизогенизация бактерий, а бактерии, содержащие профаг, называют лизогенными. Лизогенные бактериальные клетки могут обладать профагом бесконечно долгое время, при этом не лизируясь. Лизис с освобождением новых фагочастиц отмечается после воздействия на лизогенные бактерии какого-либо фактора, например УФ-излучения, которое индуцирует развитие профага в фаг. Изучение лизогенных бактерий позволило получить ряд новых данных о роли разных белков в действии фаговых генов.[ ...]
Геном хлоропластов ряда высших растений состоит из 120 генов. Хлоропластный геном очень сходен с бактериальным геномом как по организации, так и по функциям. В митохондриальном геноме человека, вероятно, отсутствуют интроны, но в ДНК хлоропластов некоторых высших растений, а также в ДНК митохондрий грибов интроны обнаружены. Считают, что хлоропластные геномы высших растений остаются без изменений примерно несколько миллионов лет. Возможно, что такая древность характерна и для митохондриальных геномов млекопитающих, включая человека.[ ...]
Современные схемы, иллюстрирующие работу генов, построены на основании логического анализа экспериментальных данных, полученных с помощью биохимических и генетических методов. Применение тонких электронно-микроскопических методов позволяет в буквальном смысле слова увидеть работу наследственного аппарата клетки. В последнее время получены электронно-микроскопические снимки, на которых видно, как на матрице бактериальной ДНК, в тех участках, где к ДНК прикреплены молекулы РНК-полимеразы (фермента, катализирующего транскрипцию ДНК в РНК), происходит синтез молекул и-РНК. Нити и-РНК, расположенные перпендикулярно к линейной молекуле ДНК, продвигаются вдоль матрицы и увеличиваются в длине. По мере удлинения нитей РНК к ним присоединяются рибосомы, которые, продвигаясь, в свою очередь, вдоль нити РНК по направлению к ДНК, ведут синтез белка.[ ...]
Трансдукция - это перенос генетического материала от бактерии-донора к бактерии-реципиенту с помощью фага. Впервые явление трансдукции было открыто в 1951 г. Ледербергом с сотрудниками у Salmonella typhimurium. Сейчас различают неспецифическую и специфическую трансдукции. При неспецифической трансдукции возможен перенос фагом любого признака от бактерии-донора к бактерии-реципиенту. Перенос осуществляется только умеренными (невирулентными) фагами. Умеренные фаги способны заражать бактерии, однако не размножаются в них и не вызывают лизиса, а включаются в ДНК бактериальной клетки и в таком неинфекционном состоянии в виде так называемого профага передаются от клетки к клетке при размножении. Культуры бактерий, содержащие профаг, называются лизогенными. В этих культурах с небольшой частотой (в одной из 102 - 105 клеток) наблюдается спонтанное размножение фага и происходит лизис клетки с освобождением фаговых частиц, обнаруживаемых с помощью бактерий-индикаторов, для которых такой фаг вирулентен.[ ...]
Опыты проводили на трехкамерной ячейке, состоящей из центральной рабочей и двух электродных камер. В рабочую камеру размером 25 X 7 X 37 мм (длина X ширина X высота), отделенную от электродных целлофановыми мембранами, помещали 750 мг ваты. Через нее снизу вверх подавали с постоянной скоростью исходный раствор исследуемых веществ. За содержанием соединений в исходных, подаваемых в рабочую камеру растворах (С0), и в растворах, выходящих из камеры (Ci), следили по максимумам поглощения белков и нуклеиновых кислот в диапазоне волновых чисел (35,5-38) X Ю3 см-1 с помощью УФ-спектрофотометра Specord UV-VIS. Электродные камеры заполняли гранулированным активированным углем и через них отдельным протоком пропускали дистиллированную воду.
2015-09-11 09:16:41
Спрашивает Мария :
Здравствуйте! Беспокоят обильные белые выделения и периодический зуд, сдала анализ на скрытые инфекции, при заборе анализа врач сказала что выделения очень обильные и есть сильное воспаление. Результаты анализа: ДНК Gardnerella vaginalis1*10^9Не более ДНК Lactobacillus spp, ДНК Atopobium vaginae2*10^9Не более ДНК Lactobacillus spp, ДНК Lactobacillus spp.8*10^7Не менее ДНК Bacteria, ДНК Bacteria (общее количество бактерий)1*10^9Не менее 10^6. Коэффициент соотношения Lactobacillus spp. к Bacteria -1.2 более "-1.0". Все остальные инфекции не обнаружены. Что это значит? Заранее большое спасибо за ответ.
Отвечает Босяк Юлия Васильевна :
Здравствуйте, Мария! У Вас банальный бактериальный вагиноз, не более. Данное заболевание провоцирует обильные выделения, которые приводят к воспалительному процессу во влагалище. Обратитесь к гинекологу для назначения терапии.
2015-05-08 20:03:38
Спрашивает наталья :
Добрый вечер. Посмотрите, пожалуйста. . РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ мазка на ИППП -- ДНК Gardnerella vaginalis 2x10^8 Копии/мл норма-- Не более ДНК Lactobacillus spp ДНК Atopobium vaginae не обнаружено норма-- Не более ДНК Lactobacillus spp ДНК Lactobacillus spp. 2x10^7 Копии/мл норма-- Не менее ДНК Bacteria ДНК Bacteria(Общее количество бактерий) 6x10^8 Копии/мл норма-- Не менее 10^6 Состояние бактериального микроценоза --- Соотношения концентраций ДНК микроорганизмов соответствуют бактериальному вагинозу.Уреа,мико,хлам,трих,гонококк,кандида не обнаружена. .В мазке на флору есть лейкоциты во влаг.14-15 ед.,шейке 18-20,уретра 3-4.кл.плоского эпителия ---единичны.гр.палочки -неб.кол-во..кокки отсутствуют. Пожалуйста, назначте лечение, уже 2 недели ничего не лечу и опять праздники. А мужу тоже нужно лечиться?Можно ли теперь жить пол. жизнью.?Очень Вас прошу помогите. Благодарю ВАС. .
Отвечает Босяк Юлия Васильевна :
Здравствуйте, Наталья! Виртуально ни один врач назначать лечение не будет. Вам необходимо с результатом анализа обратиться к гинекологу. Ничего критичного у Вас нет, банальный бактериальный вагиноз. Жить открытой половой жизнью без лечения не рекомендую.
2014-10-09 12:40:16
Спрашивает Оксана :
Добрый день.
Результаты анализов:
ДНК Gardnerella vaginalis - 7*10^7
ДНК Lactobacillus spр. - 1*10^7
ДНК Bacteria (общее количество бактерий) - 3*10^7
ДНК Ureaplasma urealiticum - 5*10^6
ДНК Staphyloccus spp.- 2*10^3
ДНК Streptococcus spp.- 1*10^4
Нужно ли делать дополнительно посевы или можно сразу идти на приём к врачу, за назначением лечения?
Отвечает Дикая Надежда Ивановна :
Оксана, добрый день! Результат обследований - информативный, необходимо лечение. Пролечитесь, не жалейте денег, присутствует комбинорованая инфекция. Удачи Вам!
2014-06-27 07:13:58
Спрашивает Инга :
Добрый день! Помогите, пожалуйста, определиться с лечением.
Сдавала мазок на пцр, выявили:
уреаплазму уреалектикум в количестве 2*10^4 (референсное значение: не обнаружено)
и микоплазму хоминис в количестве 5*10^6 (референсное значение: не обналужено).
Так же был выявлен бактериальный вагиноз:
количество лактобактерий - 1*10^5 (референсеое значение: не менее общего количества бактерий),
общее количество бактерий - 3*10^8 (референсное значение: не менее 10^6),
гарданелла вагиналис - 1*10^7 (референсное значение: не больше количества лактобактерий)
и атопобиум вагинале - 2*10^7 (референсное значение: не больше количества лактобактерий)
У меня следующий вопрос. Если я сейчас начну лечить микоплазму с уреаплазмой антибиотиком, к примеру юнидоксом, вагиноз усугубится в разы, правильно? Как быть в таком случае?
И еще вопрос по поводу лечения вагиноза. Прочитала, что атопобиум вагинае практически не реагирует на метронидазол, которым обычно лечат бактериальный вагиноз и что именно он является причиной рецедивов данного заболевания. Что бы вы посоветовали для лечения? Может есть какой-то антибиотик влияющий сразу на все: и на уреаплазму с микоплазмой и на гарданеллу с атопобиумом?
Отвечает Медицинский консультант портала «сайт» :
Добрый день, Инга! Вы мыслите в верном направлении. Антибиотики Вам не нужны, даже противопоказаны. Дело в том, что Ваш основной диагноз - урогенитальный дисбактериоз. Именно потому, что нарушился баланс нормальной микрофлоры, на слизистых начала активно размножаться условнопатогенная и транзиторная микрофлора (уреаплазма, микоплазма). Вам нужно восстановить нормальную микрофлору (препараты лактобактерий, аутовакцины, бактериофаги и т.д.), тогда и уреаплазмы с микоплазмами исчезнут с Ваших слизистых оболочек. Если проведете антибиотикотерапию, получите временное улучшение состояния, а затем дисбактериоз усугубится (так как еще больше убьете нормальную микрофлору) и неизвестно, что осядет на слизистых в следующий раз. Будьте здоровы!
2014-03-07 15:59:00
Спрашивает мария :
Здравствуйте!помогите расшифровать результаты исследования;
днк bacteria-общее количество бактерий(копий/мл)6.7*10^5
днк lactobacillus spp.(копий/мл)4.6*10^4 (не менее концентрации днк bacteria
днк gardnerella vaginalis(копий/мл)не обнаружено (не превышает концентрацию днк lactobacillus
днк atopobium vaginae(копий/мл) не обнаружено(не превышает концентрацию днк lactobacillus
заключение:снижение степени бактериальной обсемененности.
Отвечает Пурпура Роксолана Йосиповна :
По результату анализа патогенной микрофлоры не найдено, однако количество полезной микрофлоры (лактобактерий) понижено. Это может свидетельствовать о бисбактериозе влагалища.
2013-03-02 22:30:55
Спрашивает Александра :
Здравствуйте. У меня такой вопрос? Была на приеме у гинеколога, шейка матки и стенки влагалища сильно воспалены(длительное время мучают выделения, зуд) доктор взял мазок на инфекции: В мазке
ДНК Ureaplasma parvum 2*10^5
ДНК Gardnerella vaginalis Не обнаружено
ДНК Atopobium vaginae Не обнаружено
ДНК Lactobacillus spp. 1*10^7
ДНК Bacteria (общее количество бактерий) 2*10^7
1 - Коэффициент соотношения Lactobacillus spp. к Bacteria -0,1
2 - Коэффициент соотношения Lactobacillus spp. к G.vaginalis и A.vaginae 7,1
Состояние бактериального микроценоза Дисбиоз не выявлен
Что означают эти анализы?? Большое ли кол-во уреаплазмы у меня в мазке?? Дальнейший ход моих действий?? Если нужно лечение, то какое?? Спасибо
Отвечает :
Добрый день, Александра.
Уреаплазм многовато, однако, т.к. лактобактерий больше чем уреаплазм, дисбиоза пока нет, и со временем, возможно, лактобактерии их вытеснят.
Уреаплазмы относятся к группе транзиторных микроорганизмов. Другими словами, они могут временно находиться в половых путях здоровой женщины, не вызывая никаких проблем. В таком случае никакого лечения не требуется. Если же они вызывают воспаление – уреаплазмоз, нужно проводить лечение этого воспалительного процесса. Наконец, если воспаления нет, но Вы планируете, беременность или смену полового партнера, то от уреаплазм стоит избавиться.
Т.к. у вас все же есть воспаление, то лучше все же пройти курс лечения, который вам назначит ваш врач на очном приеме.
Будьте здоровы!
2013-01-14 15:36:15
Спрашивает Анна :
Помогите пожалуйста расшифровать результаты анализа. Планируем малыша.
более 50 - для мужчин
ДНК Candida albicans/glabrata/crusei (количественное исследование) Готов.
Количество ДНК Candida albicans 2*10^7 Не обнаружено Копии/мл
Количество ДНК Candida glabrata Не обнаружено Не обнаружено Копии/мл
Количество ДНК Candida crusei Не обнаружено Не обнаружено Копии/мл
ДНК U.parvum/U.urealyticum (количественное исследование) Готов.
Количество ДНК Ureaplasma parvum 4*10^6 Не обнаружено Копии/мл
Количество ДНК Ureaplasma urealyticum Не обнаружено Не обнаружено Копии/мл
Бактериальный вагиноз Готов.
ДНК Gardnerella vaginalis 3*10^8 Не более ДНК Lactobacillus spp Копии/мл
ДНК Atopobium vaginae 1*10^7 Не более ДНК Lactobacillus spp Копии/мл
ДНК Lactobacillus spp. 3*10^8 Не менее ДНК Bacteria Копии/мл
ДНК Bacteria (общее количество бактерий) 7*10^8 Не менее 10^6 Копии/мл
1 - Коэффициент соотношения Lactobacillus spp. к Bacteria -0,3 Более " - 1.0"
2 - Коэффициент соотношения Lactobacillus spp. к G.vaginalis и A.vaginae 0,0 Более "1.0"
Состояние бактериального микроценоза Мезоценоз Дисбиоз не выявлен
Исследование выполнил: Егорова Е.А.
Отвечает Серпенинова Ирина Викторовна :
Мезоценоз - это состояние микрофлоры влагалища, при котором количество лактобактерий (Lactobacterium sp) несколько ниже количества других бактерий (Bacteria), а количество гарднерелл/анаэробных бактерий (Gardnerella/ Atopobium) повышено.Иногда восстановление нормально микрофлоры происходит самостоятельно,иногда требует назначения препаратов,содержащих лактобактерии.
2012-11-30 17:53:15
Спрашивает Ольга :
Добрый день! Можете помочь расшифровать результаты анализов. О чем начать беспокоится?
Параметр Результат Референсные значения Ед.Изм.
Количество клеток в образце (параметр используется для клинической интерпретации) более 500 Более 500 - для женщин;
более 50 - для мужчин
Урогенитальные инфекции у женщин Готов.
ДНК Neisseria gonorrhoeae Не обнаружено Не обнаружено Копии/мл
ДНК Chlamydia trachomatis Не обнаружено Не обнаружено Копии/мл
ДНК Mycoplasma genitalium Не обнаружено Не обнаружено Копии/мл
ДНК Trichomonas vaginalis Не обнаружено Не обнаружено Копии/мл
ДНК Ureaplasma parvum Не обнаружено Не обнаружено Копии/мл
ДНК Ureaplasma urealyticum 1*10^4 Не обнаружено Копии/мл
ДНК Mycoplasma hominis 2*10^3 Не обнаружено Копии/мл
ДНК Candida albicans Не обнаружено Не обнаружено Копии/мл
ДНК Candida glabrata Не обнаружено Не обнаружено Копии/мл
ДНК Candida krusei Не обнаружено Не обнаружено Копии/мл
ДНК Gardnerella vaginalis 2*10^5 Не более ДНК Lactobacillus spp Копии/мл
ДНК Atopobium vaginae 1*10^4 Не более ДНК Lactobacillus spp Копии/мл
ДНК Lactobacillus spp. 2*10^7 Не менее ДНК Bacteria Копии/мл
ДНК Bacteria (общее количество бактерий) 4*10^7 Не менее 10^6 Копии/мл
1 - Коэффициент соотношения Lactobacillus spp. к Bacteria -0,2 Более " - 1.0"
2 - Коэффициент соотношения Lactobacillus spp. к G.vaginalis и A.vaginae 2,2 Более "1.0"
Состояние бактериального микроценоза Дисбиоз не выявлен Дисбиоз не выявлен
ДНК вируса простого герпеса I, II (HSV I/II), Цитомегаловируса (CMV) Готов.
Отвечает Консультант медицинской лаборатории «Синэво Украина» :
Добрый день, Ольга. В принципе, повода для беспокойства нет. Дисбиоз влагалища не выявлен. Выявленны условно-патогенные микроорганизмы, как уреаплазма, микоплазма и гарднерелла, однако их количество меньше полезной микрофлоры, поэтому дисбиоза нет. Но если вы планируете смену партнера или беременность от них все же лучше избавиться заранее. Будьте здоровы!
2012-04-27 10:28:25
Спрашивает марина :
Здравствуйте! недавно сдала анализы, получила результаты, а расшифровать не получается, помогите пожалуйста!
ДНК Gardnerella vaginalis 6*10*6,9% OKБ
ДНК Atopobium vaginae1*10*7 ,14 % OKБ
ДНК Lactobacillus sp. 2*10*7 ,менее 1% OKБ
ДНК Bacteria (общее количество бактерий) 7*10*7
ДНК Ureaplasma urealyticum ДНК не выявлена
ДНК Mycoplasma hominis 4,8*10*4
грибы рода Candida ДНК не выявлена
Результаты исследований:
ДНК N.gonorrhoeae. ДНК выявлена
ДНК C.trachomatis ДНК выявлена.
ДНК M.genitalium ДНК не выявлена
Огромное спасибо!
Отвечает Консультант медицинской лаборатории «Синэво Украина» :
Добрый день, Марина.
У Вас выявлены ДНК гонококков, хламидий, гарднерелл, микоплазм, Atopobium vaginae. А также лактобактерии – нормальная микрофлора влагалища. Так что Вам на очный прием к гинекологу для назначения курса лечения Вам и Вашему партнеру.
Будьте здоровы!
У Вас выявлены ДНК гонококков, хламидий, гарднерелл, микоплазм, Atopobium vaginae. А также лактобактерии – нормальная микрофлора влагалища. Так что Вам на очный прием к гинекологу для назначения курса лечения Вам и Вашему партнеру.
Будьте здоровы!
Популярные статьи на тему: общее количество бактерий
Среди общего числа лиц с патологией ЛОР-органов острый средний отит диагностируется примерно в 30% случаев. Течение острого среднего отита зависит от этиологии, совокупности предрасполагающих факторов, специфики морфологических проявлений.
Одним из наиболее обсуждаемых вопросов современной гастроэнтерологии (не только педиатрической) является дисбиоз кишечника, распространенность и клиническая значимость которого, по мнению отечественных педиатров, возводят это понятие в ранг серьезной...
12-13 февраля 2004 года г. Харьков принимал участников VII Международной конференции «Клиническая микробиология и антибактериальная терапия: проблемы и решения». Выступления ведущих специалистов из России, Белоруссии и Украины..
Неспецифические заболевания органов дыхания по таким показателям, как распространенность, смертность, экономические затраты занимают ведущее место в общей структуре заболеваемости населения большинства стран мира.В 2002 г. в Украине...
Новости на тему: общее количество бактерий
Здоровье человека во многом зависит от гигиены рта. Люди, у которых во рту находится большое количество микробов, чаще страдают от инфарктов, причем риск возрастает не из-за какой-то определенной бактерии, а от повышенного количества бактерий разных типов в целом. К такому выводу пришли ученые из США.
БАКТЕРИИ
обширная группа одноклеточных микроорганизмов, характеризующихся отсутствием окруженного оболочкой клеточного ядра. Вместе с тем генетический материал бактерии (дезоксирибонуклеиновая кислота, или ДНК) занимает в клетке вполне определенное место - зону, называемую нуклеоидом. Организмы с таким строением клеток называются прокариотами ("доядерными") в отличие от всех остальных - эукариот ("истинно ядерных"), ДНК которых находится в окруженном оболочкой ядре. Бактерии, ранее считавшиеся микроскопическими растениями, сейчас выделены в самостоятельное царство Monera - одно из пяти в нынешней системе классификации наряду с растениями, животными, грибами и протистами.
Ископаемые свидетельства. Вероятно, бактерии - древнейшая известная группа организмов. Слоистые каменные структуры - строматолиты, - датируемые в ряде случаев началом археозоя (архея), т.е. возникшие 3,5 млрд. лет назад, - результат жизнедеятельности бактерий, обычно фотосинтезирующих, т.н. сине-зеленых водорослей. Подобные структуры (пропитанные карбонатами бактериальные пленки) образуются и сейчас, главным образом у побережья Австралии, Багамских островов, в Калифорнийском и Персидском заливах, однако они относительно редки и не достигают крупных размеров, потому что ими питаются растительноядные организмы, например брюхоногие моллюски. В наши дни строматолиты растут в основном там, где эти животные отсутствуют из-за высокой солености воды или по другим причинам, однако до появления в ходе эволюции растительноядных форм они могли достигать огромных размеров, составляя существенный элемент океанического мелководья, сравнимый с современными коралловыми рифами.
В некоторых древних горных породах обнаружены крохотные обугленные сферы, которые также считаются остатками бактерий. Первые ядерные, т.е. эукариотические, клетки произошли от бактерий примерно 1,4 млрд. лет назад.
Экология.
Бактерий много в почве, на дне озер и океанов - повсюду, где накапливается органическое вещество. Они живут в холоде, когда столбик термометра чуть превышает нулевую отметку, и в горячих кислотных источниках с температурой выше 90° С. Некоторые бактерии переносят очень высокую соленость среды; в частности, это единственные организмы, обнаруженные в Мертвом море. В атмосфере они присутствуют в каплях воды, и их обилие там обычно коррелирует с запыленностью воздуха. Так, в городах дождевая вода содержит гораздо больше бактерий, чем в сельской местности. В холодном воздухе высокогорий и полярных областей их мало, тем не менее они встречаются даже в нижнем слое стратосферы на высоте 8 км. Густо заселен бактериями (обычно безвредными) пищеварительный тракт животных. Эксперименты показали, что для жизнедеятельности большинства видов они не обязательны, хотя и могут синтезировать некоторые витамины. Однако у жвачных (коров, антилоп, овец) и многих термитов они участвуют в переваривании растительной пищи. Кроме того, иммунная система животного, выращенного в стерильных условиях, не развивается нормально из-за отсутствия стимуляции бактериями. Нормальная бактериальная "флора" кишечника важна также для подавления попадающих туда вредных микроорганизмов.
СТРОЕНИЕ И ЖИЗНЕДЕЯТЕЛЬНОСТЬ БАКТЕРИЙ
Бактерии гораздо мельче клеток многоклеточных растений и животных. Толщина их обычно составляет 0,5-2,0 мкм, а длина - 1,0-8,0 мкм. Разглядеть некоторые формы едва позволяет разрешающая способность стандартных световых микроскопов (примерно 0,3 мкм), но известны и виды длиной более 10 мкм и шириной, также выходящей за указанные рамки, а ряд очень тонких бактерий может превышать в длину 50 мкм. На поверхности, соответствующей поставленной карандашом точке, уместится четверть миллиона средних по величине представителей этого царства.
Строение. По особенностям морфологии выделяют следующие группы бактерий: кокки (более или менее сферические), бациллы (палочки или цилиндры с закругленными концами), спириллы (жесткие спирали) и спирохеты (тонкие и гибкие волосовидные формы). Некоторые авторы склонны объединять две последние группы в одну - спириллы. Прокариоты отличаются от эукариот главным образом отсутствием оформленного ядра и наличием в типичном случае всего одной хромосомы - очень длинной кольцевой молекулы ДНК, прикрепленной в одной точке к клеточной мембране. У прокариот нет и окруженных мембранами внутриклеточных органелл, называемых митохондриями и хлоропластами. У эукариот митохондрии вырабатывают энергию в процессе дыхания, а в хлоропластах идет фотосинтез (см. также КЛЕТКА). У прокариот вся клетка целиком (и в первую очередь - клеточная мембрана) берет на себя функцию митохондрии, а у фотосинтезирующих форм - заодно и хлоропласта. Как и у эукариот, внутри бактерии находятся мелкие нуклеопротеиновые структуры - рибосомы, необходимые для синтеза белка, но они не связаны с какими-либо мембранами. За очень немногими исключениями, бактерии не способны синтезировать стеролы - важные компоненты мембран эукариотической клетки. Снаружи от клеточной мембраны большинство бактерий одето клеточной стенкой, несколько напоминающей целлюлозную стенку растительных клеток, но состоящей из других полимеров (в их состав входят не только углеводы, но и аминокислоты и специфические для бактерий вещества). Эта оболочка не дает бактериальной клетке лопнуть, когда в нее за счет осмоса поступает вода. Поверх клеточной стенки часто находится защитная слизистая капсула. Многие бактерии снабжены жгутиками, с помощью которых они активно плавают. Жгутики бактерий устроены проще и несколько иначе, чем аналогичные структуры эукариот.

"ТИПИЧНАЯ" БАКТЕРИАЛЬНАЯ КЛЕТКА
и ее основные структуры.
Сенсорные функции и поведение. Многие бактерии обладают химическими рецепторами, которые регистрируют изменения кислотности среды и концентрацию различных веществ, например сахаров, аминокислот, кислорода и диоксида углерода. Для каждого вещества существует свой тип таких "вкусовых" рецепторов, и утрата какого-то из них в результате мутации приводит к частичной "вкусовой слепоте". Многие подвижные бактерии реагируют также на колебания температуры, а фотосинтезирующие виды - на изменения освещенности. Некоторые бактерии воспринимают направление силовых линий магнитного поля, в том числе магнитного поля Земли, с помощью присутствующих в их клетках частичек магнетита (магнитного железняка - Fe3O4). В воде бактерии используют эту свою способность для того, чтобы плыть вдоль силовых линий в поисках благоприятной среды. Условные рефлексы у бактерий неизвестны, но определенного рода примитивная память у них есть. Плавая, они сравнивают воспринимаемую интенсивность стимула с ее прежним значением, т.е. определяют, стала она больше или меньше, и, исходя из этого, сохраняют направление движения или изменяют его.
Размножение и генетика. Бактерии размножаются бесполым путем: ДНК в их клетке реплицируется (удваивается), клетка делится надвое, и каждая дочерняя клетка получает по одной копии родительской ДНК. Бактериальная ДНК может передаваться и между неделящимися клетками. При этом их слияния (как у эукариот) не происходит, число особей не увеличивается, и обычно в другую клетку переносится лишь небольшая часть генома (полного набора генов), в отличие от "настоящего" полового процесса, при котором потомок получает по полному комплекту генов от каждого родителя. Такой перенос ДНК может осуществляться тремя путями. При трансформации бактерия поглощает из окружающей среды "голую" ДНК, попавшую туда при разрушении других бактерий или сознательно "подсунутую" экспериментатором. Процесс называется трансформацией, поскольку на ранних стадиях его изучения основное внимание уделялось превращению (трансформации) таким путем безвредных организмов в вирулентные. Фрагменты ДНК могут также переноситься от бактерии к бактерии особыми вирусами - бактериофагами. Это называется трансдукцией. Известен также процесс, напоминающий оплодотворение и называемый конъюгацией: бактерии соединяются друг с другом временными трубчатыми выростами (копуляционными фимбриями), через которые ДНК переходит из "мужской" клетки в "женскую". Иногда в бактерии присутствуют очень мелкие добавочные хромосомы - плазмиды, которые также могут переноситься от особи к особи. Если при этом плазмиды содержат гены, обусловливающие резистентность к антибиотикам, говорят об инфекционной резистентности. Она важна с медицинской точки зрения, поскольку может распространяться между различными видами и даже родами бактерий, в результате чего вся бактериальная флора, скажем кишечника, становится устойчивой к действию определенных лекарственных препаратов.
МЕТАБОЛИЗМ
Отчасти в силу мелких размеров бактерий интенсивность их метаболизма гораздо выше, чем у эукариот. При самых благоприятных условиях некоторые бактерии могут удваивать свою общую массу и численность примерно каждые 20 мин. Это объясняется тем, что ряд их важнейших ферментных систем функционирует с очень высокой скоростью. Так, кролику для синтеза белковой молекулы требуются считанные минуты, а бактерии - секунды. Однако в естественной среде, например в почве, большинство бактерий находится "на голодном пайке", поэтому если их клетки и делятся, то не каждые 20 мин, а раз в несколько дней.
Питание. Бактерии бывают автотрофами и гетеротрофами. Автотрофы ("сами себя питающие") не нуждаются в веществах, произведенных другими организмами. В качестве главного или единственного источника углерода они используют его диоксид (CO2). Включая CO2 и другие неорганические вещества, в частности аммиак (NH3), нитраты (NO-3) и различные соединения серы, в сложные химические реакции, они синтезируют все необходимые им биохимические продукты. Гетеротрофы ("питающиеся другим") используют в качестве основного источника углерода (некоторым видам нужен и CO2) органические (углеродсодержащие) вещества, синтезированные другими организмами, в частности сахара. Окисляясь, эти соединения поставляют энергию и молекулы, необходимые для роста и жизнедеятельности клеток. В этом смысле гетеротрофные бактерии, к которым относится подавляющее большинство прокариот, сходны с человеком.
Главные источники энергии. Если для образования (синтеза) клеточных компонентов используется в основном световая энергия (фотоны), то процесс называется фотосинтезом, а способные к нему виды - фототрофами. Фототрофные бактерии делятся на фотогетеротрофов и фотоавтотрофов в зависимости от того, какие соединения - органические или неорганические - служат для них главным источником углерода. Фотоавтотрофные цианобактерии (сине-зеленые водоросли), как и зеленые растения, за счет световой энергии расщепляют молекулы воды (H2O). При этом выделяется свободный кислород (1/2O2) и образуется водород (2H+), который, можно сказать, превращает диоксид углерода (CO2) в углеводы. У зеленых и пурпурных серных бактерий световая энергия используется для расщепления не воды, а других неорганических молекул, например сероводорода (H2S). В результате также образуется водород, восстанавливающий диоксид углерода, но кислород не выделяется. Такой фотосинтез называется аноксигенным. Фотогетеротрофные бактерии, например пурпурные несерные, используют световую энергию для получения водорода из органических веществ, в частности изопропанола, но его источником у них может служить и газообразный H2. Если основной источник энергии в клетке - окисление химических веществ, бактерии называются хемогетеротрофами или хемоавтотрофами в зависимости от того, какие молекулы служат главным источником углерода - органические или неорганические. У первых органика дает как энергию, так и углерод. Хемоавтотрофы получают энергию при окислении неорганических веществ, например водорода (до воды: 2H4 + O2 в 2H2O), железа (Fe2+ в Fe3+) или серы (2S + 3O2 + 2H2O в 2SO42- + 4H+), а углерод - из СO2. Эти организмы называют также хемолитотрофами, подчеркивая тем самым, что они "питаются" горными породами.
Дыхание. Клеточное дыхание - процесс высвобождения химической энергии, запасенной в "пищевых" молекулах, для ее дальнейшего использования в жизненно необходимых реакциях. Дыхание может быть аэробным и анаэробным. В первом случае для него необходим кислород. Он нужен для работы т.н. электронотранспортной системы: электроны переходят от одной молекулы к другой (при этом выделяется энергия) и в конечном итоге присоединяются к кислороду вместе с ионами водорода - образуется вода. Анаэробным организмам кислород не нужен, а для некоторых видов этой группы он даже ядовит. Высвобождающиеся в ходе дыхания электроны присоединяются к другим неорганическим акцепторам, например нитрату, сульфату или карбонату, или (при одной из форм такого дыхания - брожении) к определенной органической молекуле, в частности к глюкозе. См. также МЕТАБОЛИЗМ.
КЛАССИФИКАЦИЯ
У большинства организмов видом принято считать репродуктивно изолированную группу особей. В широком смысле это означает, что представители данного вида могут давать плодовитое потомство, спариваясь только с себе подобными, но не с особями других видов. Таким образом, гены конкретного вида, как правило, не выходят за его пределы. Однако у бактерий может происходить обмен генами между особями не только разных видов, но и разных родов, поэтому правомерно ли применять здесь привычные концепции эволюционного происхождения и родства, не вполне ясно. В связи с этой и другими трудностями общепринятой классификации бактерий пока не существует. Ниже приведен один из широко используемых ее вариантов.
ЦАРСТВО MONERA
Тип Gracilicutes (тонкостенные грамотрицательные бактерии)
Класс Scotobacteria (нефотосинтезирующие формы, например миксобактерии) Класс Anoxyphotobacteria (не выделяющие кислорода фотосинтезирующие формы, например пурпурные серные бактерии) Класс Oxyphotobacteria (выделяющие кислород фотосинтезирующие формы, например цианобактерии)
Тип Firmicutes (толстостенные грамположительные бактерии)
Класс Firmibacteria (формы с жесткой клеткой, например клостридии)
Класс Thallobacteria (разветвленные формы, например актиномицеты)
Тип Tenericutes (грамотрицательные бактерии без клеточной стенки)
Класс Mollicutes (формы с мягкой клеткой, например микоплазмы)
Тип Mendosicutes (бактерии с неполноценной клеточной стенкой)
Класс Archaebacteria (древние формы, например метанобразующие)
Домены. Недавние биохимические исследования показали, что все прокариоты четко разделяются на две категории: маленькую группу архебактерий (Archaebacteria - "древние бактерии") и всех остальных, называемых эубактериями (Eubacteria - "истинные бактерии"). Считается, что архебактерии по сравнению с эубактериями примитивнее и ближе к общему предку прокариот и эукариот. От прочих бактерий они отличаются несколькими существенными признаками, включая состав молекул рибосомной РНК (pРНК), участвующей в синтезе белка, химическую структуру липидов (жироподобных веществ) и присутствие в клеточной стенке вместо белково-углеводного полимера муреина некоторых других веществ. В приведенной выше системе классификации архебактерии считаются лишь одним из типов того же царства, которое объединяет и всех эубактерий. Однако, по мнению некоторых биологов, различия между архебактериями и эубактериями настолько глубоки, что правильнее рассматривать архебактерии в составе Monera как особое подцарство. В последнее время появилось еще более радикальное предложение. Молекулярный анализ выявил между двумя этими группами прокариот столь существенные различия в структуре генов, что присутствие их в рамках одного царства организмов некоторые считают нелогичным. В связи с этим предложено создать таксономическую категорию (таксон) еще более высокого ранга, назвав ее доменом, и разделить все живое на три домена - Eucarya (эукариоты), Archaea (архебактерии) и Bacteria (нынешние эубактерии).
ЭКОЛОГИЯ
Две важнейшие экологические функции бактерий - фиксация азота и минерализация органических остатков.
Азотфиксация. Связывание молекулярного азота (N2) с образованием аммиака (NH3) называется азотфиксацией, а окисление последнего до нитрита (NO-2) и нитрата (NO-3) - нитрификацией. Это жизненно важные для биосферы процессы, поскольку растениям необходим азот, но усваивать они могут лишь его связанные формы. В настоящее время примерно 90% (ок. 90 млн. т) годового количества такого "фиксированного" азота дают бактерии. Остальное количество производится химическими комбинатами или возникает при разрядах молний. Азот воздуха, составляющий ок. 80% атмосферы, связывается в основном грамотрицательным родом ризобиум (Rhizobium) и цианобактериями. Виды ризобиума вступают в симбиоз примерно с 14 000 видов бобовых растений (семейство Leguminosae), к которым относятся, например, клевер, люцерна, соя и горох. Эти бактерии живут в т.н. клубеньках - вздутиях, образующихся на корнях в их присутствии. Из растения бактерии получают органические вещества (питание), а взамен снабжают хозяина связанным азотом. За год таким способом фиксируется до 225 кг азота на гектар. В симбиоз с другими азотфиксирующими бактериями вступают и небобовые растения, например ольха. Цианобактерии фотосинтезируют, как зеленые растения, с выделением кислорода. Многие из них способны также фиксировать атмосферный азот, потребляемый затем растениями и в конечном итоге животными. Эти прокариоты служат важным источником связанного азота почвы в целом и рисовых чеков на Востоке в частности, а также главным его поставщиком для океанских экосистем.
Минерализация. Так называется разложение органических остатков до диоксида углерода (CO2), воды (H2O) и минеральных солей. С химической точки зрения, этот процесс эквивалентен горению, поэтому он требует большого количества кислорода. В верхнем слое почвы содержится от 100 000 до 1 млрд. бактерий на 1 г, т.е. примерно 2 т на гектар. Обычно все органические остатки, попав в землю, быстро окисляются бактериями и грибами. Более устойчиво к разложению буроватое органическое вещество, называемое гуминовой кислотой и образующееся в основном из содержащегося в древесине лигнина. Оно накапливается в почве и улучшает ее свойства.
БАКТЕРИИ И ПРОМЫШЛЕННОСТЬ
Учитывая разнообразие катализируемых бактериями химических реакций, неудивительно, что они широко используются в производстве, в ряде случаев с глубокой древности. Славу таких микроскопических помощников человека прокариоты делят с грибами, в первую очередь - дрожжами, которые обеспечивают большую часть процессов спиртового брожения, например при изготовлении вина и пива. Сейчас, когда стало возможным вводить в бактерии полезные гены, заставляя их синтезировать ценные вещества, например инсулин, промышленное применение этих живых лабораторий получило новый мощный стимул. См. также ГЕННАЯ ИНЖЕНЕРИЯ.
Пищевая промышленность. В настоящее время бактерии применяются этой отраслью в основном для производства сыров, других кисломолочных продуктов и уксуса. Главные химические реакции здесь - образование кислот. Так, при получении уксуса бактерии рода Acetobacter окисляют этиловый спирт, содержащийся в сидре или других жидкостях, до уксусной кислоты. Аналогичные процессы происходят при квашении капусты: анаэробные бактерии сбраживают содержащиеся в листьях этого растения сахара до молочной кислоты, а также уксусной кислоты и различных спиртов.
Выщелачивание руд. Бактерии применяются для выщелачивания бедных руд, т.е. переведения из них в раствор солей ценных металлов, в первую очередь меди (Cu) и урана (U). Пример - переработка халькопирита, или медного колчедана (CuFeS2). Кучи этой руды периодически поливают водой, в которой присутствуют хемолитотрофные бактерии рода Thiobacillus. В процессе своей жизнедеятельности они окисляют серу (S), образуя растворимые сульфаты меди и железа: CuFeS2 + 4O2 в CuSO4 + FeSO4. Такие технологии значительно упрощают получение из руд ценных металлов; в принципе, они эквивалентны процессам, протекающим в природе при выветривании горных пород.
Переработка отходов. Бактерии служат также для превращения отходов, например сточных вод, в менее опасные или даже полезные продукты. Сточные воды - одна из острых проблем современного человечества. Их полная минерализация требует огромных количеств кислорода, и в обычных водоемах, куда принято сбрасывать эти отходы, его для их "обезвреживания" уже не хватает. Решение заключается в дополнительной аэрации стоков в специальных бассейнах (аэротенках): в результате бактериям-минерализаторам хватает кислорода для полного разложения органики, и одним из конечных продуктов процесса в наиболее благоприятных случаях становится питьевая вода. Остающийся по ходу дела нерастворимый осадок можно подвергнуть анаэробному брожению. Чтобы такие водоочистные установки отнимали как можно меньше места и денег, необходимо хорошее знание бактериологии.
Другие пути использования. К другим важным областям промышленного применения бактерий относится, например, мочка льна, т.е. отделение его прядильных волокон от других частей растения, а также производство антибиотиков, в частности стрептомицина (бактериями рода Streptomyces).
БОРЬБА С БАКТЕРИЯМИ В ПРОМЫШЛЕННОСТИ
Бактерии приносят не только пользу; борьба с их массовым размножением, например в пищевых продуктах или в водных системах целлюлозно-бумажных предприятий, превратилась в целое направление деятельности. Пища портится под действием бактерий, грибов и собственных вызывающих автолиз ("самопереваривание") ферментов, если не инактивировать их нагреванием или другими способами. Поскольку главная причина порчи все-таки бактерии, разработка систем эффективного хранения продовольствия требует знания пределов выносливости этих микроорганизмов. Одна из наиболее распространенных технологий - пастеризация молока, убивающая бактерии, которые вызывают, например, туберкулез и бруцеллез. Молоко выдерживают при 61-63° С в течение 30 мин или при 72-73° С всего 15 с. Это не ухудшает вкуса продукта, но инактивирует болезнетворные бактерии. Пастеризовать можно также вино, пиво и фруктовые соки. Давно известна польза хранения пищевых продуктов на холоде. Низкие температуры не убивают бактерий, но не дают им расти и размножаться. Правда, при замораживании, например, до -25° С численность бактерий через несколько месяцев снижается, однако большое количество этих микроорганизмов все же выживает. При температуре чуть ниже нуля бактерии продолжают размножаться, но очень медленно. Их жизнеспособные культуры можно хранить почти бесконечно долго после лиофилизации (замораживания - высушивания) в среде, содержащей белок, например в сыворотке крови. К другим известным методам хранения пищевых продуктов относятся высушивание (вяление и копчение), добавка больших количеств соли или сахара, что физиологически эквивалентно обезвоживанию, и маринование, т.е. помещение в концентрированный раствор кислоты. При кислотности среды, соответствующей pH 4 и ниже, жизнедеятельность бактерий обычно сильно тормозится или прекращается.
БАКТЕРИИ И БОЛЕЗНИ
ИЗУЧЕНИЕ БАКТЕРИЙ
Многие бактерии нетрудно выращивать в т.н. культуральной среде, в состав которой могут входить мясной бульон, частично переваренный белок, соли, декстроза, цельная кровь, ее сыворотка и другие компоненты. Концентрация бактерий в таких условиях обычно достигает примерно миллиарда на кубический сантиметр, в результате чего среда становится мутной. Для изучения бактерий необходимо уметь получать их чистые культуры, или клоны, представляющие собой потомство одной-единственной клетки. Это нужно, например, для определения того, какой вид бактерии инфицировал больного и к какому антибиотику данный вид чувствителен. Микробиологические образцы, например, взятые из горла или ран мазки, пробы крови, воды или других материалов, сильно разводят и наносят на поверхность полутвердой среды: на ней из отдельных клеток развиваются округлые колонии. Отверждающим культуральную среду агентом обычно служит агар - полисахарид, получаемый из некоторых морских водорослей и почти ни одним видом бактерий не перевариваемый. Агаровые среды используют в виде "косячков", т.е. наклонных поверхностей, образующихся в стоящих под большим углом пробирках при застывании расплавленной культуральной среды, или в виде тонких слоев в стеклянных чашках Петри - плоских круглых сосудах, закрываемых такой же по форме, но чуть большей по диаметру крышкой. Обычно через сутки бактериальная клетка успевает размножиться настолько, что образует легко заметную невооруженным глазом колонию. Ее можно перенести на другую среду для дальнейшего изучения. Все культуральные среды должны быть перед началом выращивания бактерий стерильными, а в дальнейшем следует принимать меры против поселения на них нежелательных микроорганизмов. Чтобы рассмотреть выращенные таким способом бактерии, прокаливают на пламени тонкую проволочную петлю, прикасаются ею сначала к колонии или мазку, а затем - к капле воды, нанесенной на предметное стекло. Равномерно распределив взятый материал в этой воде, стекло высушивают и два-три раза быстро проводят над пламенем горелки (сторона с бактериями должна быть обращена вверх): в результате микроорганизмы, не повреждаясь, прочно прикрепляются к субстрату. На поверхность препарата капают краситель, затем стекло промывают в воде и вновь сушат. Теперь можно рассматривать образец под микроскопом. Чистые культуры бактерий идентифицируют главным образом по их биохимическим признакам, т.е. определяют, образуют ли они из определенных сахаров газ или кислоты, способны ли переваривать белок (разжижать желатину), нуждаются ли для роста в кислороде и т.д. Проверяют также, окрашиваются ли они специфическими красителями. Чувствительность к тем или иным лекарственным препаратам, например антибиотикам, можно выяснить, поместив на засеянную бактериями поверхность маленькие диски из фильтровальной бумаги, пропитанные данными веществами. Если какое-либо химическое соединение убивает бактерии, вокруг соответствующего диска образуется свободная от них зона.
Энциклопедия Кольера. - Открытое общество . 2000 .
Основной секрет органической жизни кроется в способности к размножению и передаче наследственной информации от предыдущих поколений потомкам через довольно простой механизм самокопирования макромолекулы ДНК каждой живой клетки. Каждой, независимо от того, состоит организм из большого количества клеток или же речь идет о тех ДНК, которые находятся в клетках бактерий, этих одноклеточных простейших организмов, не всегда способных даже в большую колонию собраться.
Как у всех представителей органической жизни, наследственная (генетическая) информация бактерий хранится в их ДНК. Что такое генетическая информация? Какая структура хранит наследственную информацию?
- Генетическая информация – это определенная последовательность нуклеотидов. Другого секрета в ядре нет. Копируя эту последовательность, клетка синтезирует самые разнообразные белки. Они же решают все остальные вопросы организма, начиная с организационных, заканчивая снабжением клетки строительным материалом.
- Макромолекула ДНК – четыре нуклеиновых основания (аденин, гуанин, тимин и цитозин), объединенные в двойную спираль сахаром дезоксирибозой и остатками фосфорной кислоты. Именно нуклеиновые основания кодируют последовательность сборки белков независимо от того, есть оформленное ядро в клетке или нет.
Дезоксирибонуклеиновая кислота бактерий имеет такое же строение, как молекулы – хранители наследственной информации всех остальных живых существ на планете. Так же, как все другие органические клетки, бактерия образует из ДНК хромосомы. Но это не значит, что других отличий нет.
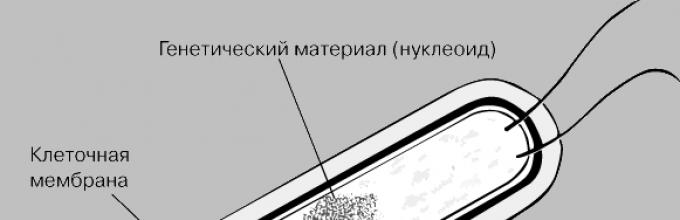
Фундаментальным отличием бактерии является то, что у нее нет клеточного ядра, наследственная информация бактерии не собрана в клеточное ядро, это просто кольцевая молекула, которая прилеплена к одной из стенок цитоплазматической мембраны.
 Однако то обстоятельство, что ядра нет, не препятствует активным процессам репликации и трансляции с использованием этого хранителя наследственной информации. Чтобы понять, как происходит передача информации, нужно понимать, что такое хромосомы, гены и клеточное ядро.
Однако то обстоятельство, что ядра нет, не препятствует активным процессам репликации и трансляции с использованием этого хранителя наследственной информации. Чтобы понять, как происходит передача информации, нужно понимать, что такое хромосомы, гены и клеточное ядро.
- Ген – участок макромолекулы, на котором записана последовательность нуклеотидов, позволяющая собирать один определенный вид белка. Другой информации в генах нет.
- Хромосома – комбинация цепи ДНК с белками гистонами, которые ее структурируют и придают ей определенную форму перед тем, как клетка начинает делиться. В фазе, когда деление не происходит, в клетке (или в ядре, если речь идет о ядерных эукариотах) как таковых хромосом нет.
- Клеточное ядро – это клеточная структура, которая содержит наследственную информацию, структурированную в хромосому, когда клетка готовится к делению. В ней инициируется сам процесс деления. Важно помнить, что у бактерий клеточного ядра нет.
Если в эукариотической клетке при делении используются обособленные, специально формирующиеся для удобства деления структуры, то как же происходит размножение бактерий в условиях неоформленного кажущегося сумбура в отсутствие клеточного ядра?
Дезоксирибонуклеиновая кислота бактериальной клетки
Бактериальная молекула ДНК хоть изображается как кольцевая довольно объемная структура, которая располагается в центре клетки, на самом деле представляет собой довольно компактное образование, локализованное на ограниченных участках цитоплазмы.
 Ввиду отсутствия ядерной мембраны, которая бы отгораживала скомпонованную бактериальную макромолекулу от других клеточных структур, генетический аппарат безъядерных организмов нельзя ассоциировать с генетическим аппаратом эукариотов, поэтому генетический аппарат прокариотов назвали нуклеоид.
Ввиду отсутствия ядерной мембраны, которая бы отгораживала скомпонованную бактериальную макромолекулу от других клеточных структур, генетический аппарат безъядерных организмов нельзя ассоциировать с генетическим аппаратом эукариотов, поэтому генетический аппарат прокариотов назвали нуклеоид.
Характерные черты нуклеоида:
- ДНК, в которой содержится нескольких тысяч генов.
- Гены расположены линейно и называются хромосомой. Хромосома бактерии – это линейная совокупность ее генов.
- Макромолекула также сворачивается белками, похожими на эукариотические гистоны.
Нуклеоид крепится к цитоплазматической мембране в тех точках, где начинается и заканчивается репликация (самокопирование).

Экспериментальным путем установлено, что нуклеоид и хромосома – это не одно и то же. Увеличение количества хромосом (линейных генов) – свидетельство того, что бактерии активно делятся. Один нуклеоид может состоять из одной хромосомы или нескольких ее копий. Так, в период деления азотобактерия реплицируется до 20-25 хромосом (копий нуклеоида).
Процесс копирования
В теоретических конструкциях, разработанных микробиологами в те годы, когда изучать сложные молекулярные процессы экспериментальным путем было очень сложно или практически невозможно, копирование дезоксирибонуклеиновой кислоты может осуществляться тремя способами:
- Консервативный, при котором двойная родительская спираль не раскручивается, а двойная дочерняя спираль полностью образовывается из нового материала.
- Дисперсивный, при котором родительская макромолекула распадается на фрагменты, а дочерние формируются на нуклеотидных последовательностях этих фрагментов как на матрицах.
- Полуконсервативный. Согласно этой модели, двойная спираль раскручивается, и каждая цепь спирали служит матрицей для дочерних ДНК. Формируется так называемый гибрид старой макромолекулы и цепи, созданной из новых компонентов.
Когда в 1957 году был найден способ отслеживания процессов, происходящих в бактериальной ДНК при ее репликации, было установлено, что дезоксирибонуклеиновая кислота реплицируется полуконсервативным путем, то есть через раскручивание и использование раскрученных участков в качестве матриц для синтеза новых макромолекул.

Сам процесс репликации бактериальной ДНК очень схож с репликацией ДНК остальных органических механизмов. Происходит он по следующей схеме:
- ДНК-хеликазы раскручивают и разрывают двойную спираль, двигаясь вдоль сахарофосфатного остова дезоксирибонуклеиновой кислоты.
- Ферменты полимеразы катализируют реакции присоединения к однонитевым фрагментам дезоксирибонуклеиновой кислоты комплиментарных нуклеиновых оснований.
После репликации происходит удвоение всех основных частей клетки: органелл, цитоплазматической мембраны, клеточной стенки, и бактериальная клетка распадается надвое.
Проблематика
Помимо исключительно научного интереса в изучении ДНК бактерий, механизм репликации и передачи наследственной информации от одной клетки к другой также имеют исключительную практическую важность.
Широко известный факт, что бактерии очень быстро адаптируются при воздействии на них антибиотиков и начинают выработку определенных белков-антител, которые блокируют разрушительное действие антибиотических средств на клетку бактерии. В следующих поколениях бактерий эта устойчивость к конкретной группе антибактериальных препаратов сохраняется.
 Более того, благодаря горизонтальному переносу генов (не в процессе деления, а в процессе простого контакта одной бактерии с другой) такая генетическая информация также передается, делая устойчивыми к антибиотикам все большее количество видов бактерий.
Более того, благодаря горизонтальному переносу генов (не в процессе деления, а в процессе простого контакта одной бактерии с другой) такая генетическая информация также передается, делая устойчивыми к антибиотикам все большее количество видов бактерий.
Изучением этих свойств бактерий, определением того, как посторонний ген включается в общую структуру дезоксирибонуклеиновой кислоты, и занимается современная микробиология.







